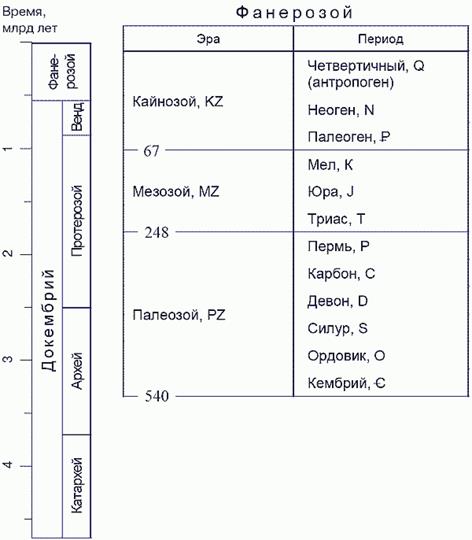
2. ГЕОХРОНОЛОГИЯ И ДВИЖЕНИЯ ЗЕМНОЙ КОРЫ
Задание 2.1
Общие особенности мезозойской эры
Мезозойская эра - так называется в геологии весьма значительный период в истории развития Земли, следовавший за палеозойской эрой и предшествовавший кайнозойской эре, к которой геологи относят и переживаемый нами период. Отложения М. эры составляют М. группу слоев, которая, в свою очередь, распадается на три крупных отдела, или системы: триасовую, или триас, юрскую и меловую. Общая мощность слоев М. группы определяется для некоторых местностей Европы по крайней мере в 3300 м, а площадь, занятая в настоящее время М. образованиями на земной поверхности, равняется, по Тилло, 20 млн. кв. км, что составляет почти 1 /5 часть всей суши. В Европе, где М. эра впервые изучена и установлена, начало ее знаменуется значительным ослаблением вулканической деятельности, столь энергично проявлявшейся в палеозойскую эру. Период затишья продолжается в течение всей М. эры и нарушается лишь с наступлением кайнозойской. Сравнительно небольшие извержения порфиров происходили лишь в сев. Шотландии и Тироле, а может быть, к этому же времени относятся некоторые изверженные породы Крыма и Кавказа. Напластование осадочных пород М. группы является по большей части спокойным и мало нарушенным, особенно по сравнению с пластами палеозойской группы; они выведены из горизонтального положения и собраны в крутые складки только в высоких горных хребтах, каковы: Юра, Альпы, Крым и Кавказ и в ближайшем соседстве с некоторыми другими горными массивами. Осадочные образования М. группы отличаются от предшествовавших палеозойских и по петрографическому составу. Известняки пользуются здесь гораздо большим распространением, чем в пластах палеозойской группы; напротив, глинистые, кремнистые сланцы и кварциты совсем не встречаются - место их занимают глины, мергели и песчаники, часто весьма рыхлые. В органическом мире также происходят чрезвычайно существенные изменения. В середине эры появляются первые млекопитающие, из отряда сумчатых, костистые рыбы, а в конце эры первые птицы, правда, еще сохранившие некоторые особенности организации пресмыкающихся, и, наконец, лиственные деревья. Чрезвычайным развитием пользовались в это время, особенно характерные для М. эры, разнообразные морские и наземные ящеры, иногда гигантских размеров, а из моллюсков - аммониты и белемниты. Плеченогие, столь распространенные в палеозойских отложениях, уступают место устрицам и другим пластинчатожаберным, морские лилии уступают преобладание морским ежам, кораллы четверного палеозойского типа сменяются шестерными (Hexacorallia). Окончательно вымирают к этому времени трилобиты и цистидеи. В мире растений лепидодендроны, каламиты и другие древовидные тайнобрачные палеозойской эры сменяются саговыми и хвойными деревьями.
Задание 2.2
Триас — сравнительно короткий период, он начался 230 миллионов лет назад и кончился 195 миллионов лет назад. В триасе на больших территориях распространяется засушливый климат, вызывающий вымирание разных групп амфибий и развитие разнообразных рептилий.
Некоторые рептилии становятся хищными и нападают на себе подобных, другие — типичными травоядными, третьи возвращаются жить в моря, где для них было много пищи (головоногих моллюсков и разнообразных рыб).
Из триасовых рептилий до наших дней дожила знаменитая гаттерия — живое ископаемое, населяющее несколько островков у берегов Австралии. Мало изменились с тех времен и черепахи.
Считается, что в триасе появились первые млекопитающие . Они были размером с крысу и были похожи на опоссума — сумчатую крысу, которая и сейчас живет в Центральной Америке. По нашим условным часам это произошло примерно за полчаса до полудня. В триасе продолжается начавшееся в перми вымирание гигантских споровых растений. Все большее распространение получают голосеменные.
Триасовое развитие характеризуется четкими минимумами в начале и конце периода, совпадающими с минимумами морского разнообразия. Максимум между ними у неморских животных смещен ближе к концу периода. Уменьшение общего разнообразия в позднем триасе определяется уменьшением числа семейств рыб, тетрапод и членистоногих (без насекомых), тогда как разнообразие моллюсков и водных насекомых в позднем триасе несколько увеличивается. В раннем и среднем триасе диверсификация всех изученных крупных групп происходит согласованно, лишь у насекомых она выражена -менее отчетливо.
Юра. Разнообразие монотонно увеличивается в течение всего периода, в средней юре медленнее, чем в ранней и особенно в поздней. В море разнообразие также постоянно нарастает, но более равномерно. При анализе изменений числа семейств в крупных группах обращает на себя внимание сохранение в ранней и средней юре стабильно низкого уровня разнообразия рыб, тетрапод, артропод (кроме насекомых) и моллюсков. У моллюсков оно даже несколько снижается по сравнению с концом триаса (рис. 2, о). У рыб и тетрапод стабильный раннесреднеюрский уровень составляет около 1/3 наибольшего разнообразия в перми. Для перечисленных групп характерна также низкая скорость появления семейств. Так, скорость появления рыб и водных тетрапод в ранней и средней юре составляет 0,15 семейства за 1 млн. лет, тогда как в поздней юре семейства тетрапод появлялись со скоростью 0,60, как и рыбы в триасе (0,62). Соответствующие относительные скорости составляют 0,022—0,024 против 0,058—0,065 млн. лет-1 . Простейшее объяснение этой странной, не имеющей аналогов эпохи застоя может состоять в сужении жизненного пространства биоты внутриконтинентальных водоемов в талассокрспическую эпоху, сходно с тем, как описывается моделями островной биогеографии. Раннеюрский рост общего разнообразия определяется в основном диверсификацией водных насекомых, тогда как в поздней юре происходит более активный рост разнообразия также водных тетрапод, в меньшей мере — моллюсков и членистоногих (без насекомых).
Ордовик. Первое зарегистрированное в палеонтологической летописи вхождение животных в биоту внутриконтинентальных водоемов, по данным Г. Греффина, относится к аренигу, когда появились пресноводные представители бесчелюстных семейства Arandaspididae. В лландейло к ним добавились представители семейств Astraspididae и Eriptichiidae. Только последние из них переходят в силурийскую внутриконтинентальную водную биоту, в связи, с чем разнообразие семейств снижается в позднем ордовике с двух до одного, и появляется слабо выраженный минимум на границе ордовика и силура. Он соответствует минимуму в разнообразии морских животных, но материал по неморским слишком скуден.
Силур. В силуре общее разнообразие семейств формируется позвоночными (почти целиком бесчелюстными) и членистоногими (почти целиком хелицеровыми), вклад которых почти равен. Обе крупные группы в течение почти всего периода увеличивают свое разнообразие, однако в конце его ведут себя по-разному. Разнообразие членистоногих продолжает увеличиваться до границы с девоном, тогда как у позвоночных оно несколько уменьшается в пржидоле. Суммарная кривая на пржидольском участке становится более пологой и характеризует замедление роста общего разнообразия, отличаясь от кривой для морских животных, на которой границе с девоном соответствует отчетливый, хотя и не глубокий, минимум
Задание 2.3
Разрывные дислокации образуются в результате интенсивных тектонических движений, сопровождающиеся разрывом сплошности пород и смещением слоев относительно друг друга. Амплитуда смещения может быть от нескольких сантиметров до километров при ширине трещин до нескольких метров. К разрывным дислокациям относятся сбросы, взбросы, грабены, горсты, сдвиги и надвиги (рис.: а – неподвижная часть земной коры, б – подвижная часть).
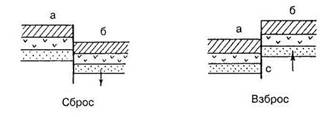
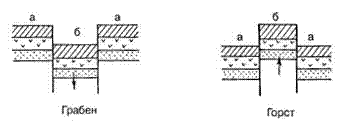
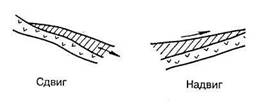
Разрывные типы дислокаций
Сбросы – разрывные нарушения, когда подвижная часть земной коры опустилась вниз по отношению к неподвижной.
Взброс – разрывное нарушение, когда подвижная часть земной коры поднялась в результате тектонического движения по отношению к неподвижной.
Грабен – когда подвижный участок земной коры опустился по отношению к двум неподвижным участкам в результате тектонического движения.
Горст – обратное грабену движение.
Сдвиг – представляет собой разрывное нарушение, в котором происходит горизонтальное смещение горных пород по простиранию.
Надвиг – обратное сдвигу перемещение.
С инженерно-геологической точки зрения наиболее благоприятными местами строительства являются горизонтальное залегание горных пород, где присутствует большая их мощность, однородность состава. Фундаменты зданий и сооружений располагаются в однородной грунтовой среде, при этом создается равномерная сжимаемость слоев под весом сооружения и создается наибольшая их устойчивость
Наличие дислокации резко изменяет и усложняет инженерно-геологические условия строительства – нарушается однородность грунтов основания фундамента сооружений, образуются зоны дробления (разрывы), снижается прочность пород, по трещинам разрывов происходят смещения, нарушается режим подземных вод. Это вызывает неравномерную сжимаемость грунтов и деформацию самого сооружения вследствие неравномерной осадки различных его частей.
Задание 2.4
Грабен
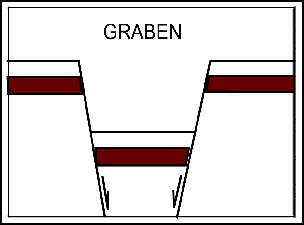 Гра
беном
(нем. "graben" - копать
) называется структура, ограниченная с двух сторон сбросами
Гра
беном
(нем. "graben" - копать
) называется структура, ограниченная с двух сторон сбросами
Строение грабена замаскировано гравитационными нормальными сбросами, представляющими побочное явление, сопровождающее взбросы
Совершенно своеобразный тектонический тип представляют узкие впадины проседания типа Байкала, Осло, верхней долины Рейна, Восточноафриканских грабенов. На единство этих геологических структур указал Н.С. Шатский (1932).
Первоначальные представления об образовании Байкальской впадины были высказаны еще П. Далласом, считавшим, что одновременно с поднятием берегов Байкала произошло оседание его дна. И.Д. Черский (1886) считал, что впадина, занятая Байкалом, представляет крутую синклинальную складку, возникшую в раннем палеозое или древнее. В.А. Обручев в 1897 г. пришел к выводу, что впадина Байкала вместе с целой системой забайкальских депрессий представляет сложную цепь грабенов, образовавшихся при расколах жесткой глыбы байкальского кристаллического массива. При этом он рассматривал впадину Байкала не как изолированное образование, а как наиболее крупный грабен среди целой системы впадин, развитых на большой территории. Изучавший Африканские грабены Дж. Грегори (1921) и другие исследователи также пришли к выводу об образовании узких впадин путем обрушения земной коры по параллельным сбросам. Г. Клоос (1939) связал грабены с формированием сводовых поднятий и обрушением замковых частей сводов. В целом, гипотезы, предполагающие образование узких впадин в связи с растяжением земной коры и обрушением, получили название гипотез рифта (rift—расселина ). Э. Вейланд (1930, 1933, 1934) и Виллис (1934) для объяснения способа образования впадин в противовес рифтовой гипотезе предложили так называемую гипотезу рампа (ramp—взброс ). Согласно последней гипотезе, грабены образованы в условиях сжатия, поднявшего висячие бока надвигавшихся глыб. Глыба, находившаяся в лежачем боку, опускалась под воздействием надвигавшегося блока. Механизм образования грабена в последнем случае представляется примерно так. Тангенциальное тектоническое усилие действует в одном направлении, со стороны активной глыбы. Под воздействием этой глыбы блок, находящийся в лежачем боку надвига, не только погружается, но и пододвигается под противостоящий, пассивный блок, поэтому возникает плоскость разрыва, параллельная надвигу, но падающая в противоположном направлении. В связи с пододвиганием блока лежачего бока происходит также поднятие противостоящей, пассивной глыбы, составляющей висячий бок надвига.
Задание 2.5
1. Зная период Т и амплитуду А колебаний сейсмической волнывычислим сейсмическое ускорение α и коэффициент сейсмичности Кs:
Где g=9800 мм/с².
α=;
![]()
QUOTE
29-04-2015, 00:58