ЗМІСТ
ВСТУП
РОЗДІЛ 1. ОСОБЛИВОСТІ МОРФО-ЦИТОЛОГІЧНОЇ ОРГАНІЗАЦІЇ МІКРООРГАНІЗМІВ
РОЗДІЛ 2. МОЖЛИВОСТІ ВИКОРИСТАННЯ МІКРОБІОЛОГІЧНИХ ПРЕПАРАТІВ ДЛЯ БОРОТЬБИ ІЗ ШКІДНИКАМИ
2.1 Мікробіологічні інсектициди
2.2 Інтродукція мікроорганізмів та мікробіологічний метод боротьби
2.3 Нешкідливість мікробіологічних агентів
РОЗДІЛ 3. МІКРООРГАНІЗМИ ЯК ДЖЕРЕЛО НОВИХ ІНСЕКТИЦИДНИХ ПРЕПАРАТІВ
3.1 Класифікація бактеріальних токсинів
3.2 Класифікація за анатомічним розміщенням
3.3 Класифікація за місцем або механізмом дії
3.4 Класифікація за структурою молекули токсину
3.5 Проблема токсинів у патології комах
3.6 Практичні можливості і перспективи використання інсектицидів
ВИСНОВОК
ЛІТЕРАТУРА
ВСТУП
Матеріали багатьох досліджень говорять про цінності мікробіологічного методу як практичної зброї проти деяких шкідливих комах. Вони виявляють також проведення активних досліджень фундаментальних і практичних сторін патології комах, що повинно гарантувати подальший прогрес. Патогени комах, застосовуваних повторно, позначають терміном "мікробіологічні інсектициди", патогени, які здатні виживати і поширюватися в популяціях шкідливих комах, розглядають як "інтродукції". Ефективність біологічного методу боротьби зі шкідниками рослин подекуди буває досить високою. Зафіксовано випадки, коли застосування мікробних препаратів викликало масову загибель шкідників. Проте результати застосування мікробіологічного методу значною мірою залежать від кліматичних умов, стану шкідників та інших чинників.
Збудниками інфекційних захворювань шкідливих комах можуть бути також гриби і віруси.В різних країнах світу на основі патогенних мікроорганізмів випускаються понад 50 препаратів для боротьби з 200 видами шкідливих комах. Найкраще налагоджено виробництво таких бактеріальних препаратів: ентобактерин, дендробацилін, ліпідоцид, а з грибкових препаратів — актинін, боверин та ін.
Мета роботи – розглянути можливості використання мікроорганізмів з метою створення екологічно безпечних антимікробних засобів.
Завданнями роботи є:
1) розглянути особливості морфо-цитологічної будови мікроорганізмів;
2) охарактеризувати можливості використання мікробіологічних препаратів для боротьби із шкідниками;
3) дати характеристику мікроорганізмам як джерелу нових інсектицидних препаратів.
РОЗДІЛ 1. ОСОБЛИВОСТІ МОРФО-ЦИТОЛОГІЧНОЇ ОРГАНІЗАЦІЇ МІКРООРГАНІЗМІВ
Ультраструктура прокаріотної клітини вивчаються за допомогою електронно-мікроскопічних, мікрохімічних та інших методів, які дають змогу з високою точністю визначити будову і хімічний склад бактерій. Завдяки цим методам вдалося встановити, що прокаріотна клітина має низку принципових особливостей ультраструктурної і хімічної організації порівняно з еукаріотною клітиною (рис. 1).
Зовні цитоплазматичної мембрани бактеріальної клітини розміщуються так звані поверхневі структури: оболонка, капсула, слизовий чохол, джгутики і ворсинки (війки). Цитоплазматичну мембрану разом з цитоплазмою, органелами і включеннями прийнято називати протопластом. Розглянемо спочатку будову, хімічний склад і функції поверхневих структур бактеріальної клітини.

Рис. 1. Комбіноване схематичне зображення прокаріотпої клітини (за Г. Шлегелем, 1972):
І — поверхневі структури: 1 — оболонка клітини; 2 — капсула; 3 — слизові виділення; 4 — слизовий чохол; 5 — джгутики; 6 — війки.
ІІ — цитоплазматичні клітинні структури: 7 — цитоплазматична мембрана; 8 — нуклеоїд; 9 — рибосоми; 10 — цитоплазма; 11 — хроматофори; 72 — хлоросоми; ЇЗ — пластинчасті тилакоїди; 14 — фікобілісоми; 15 — трубчасті тилакоїди; 16 — мезосома; 17 — аеросома (газова вакуоля); 18 — ламелярні структури.
ІІ — записні речовини: 19 — полісахаридні гранули; 20— гранули полі-(3-окси масляної кислоти; 21 — гранули поліфосфату; 22— гранули ціанофіцину; 23 — карбоксисоми; 24— включення сірки; 25 — крапельки жиру; 26 — гранули вуглеводів
За будовою і хімічним складом оболонка бактеріальної клітини суттєво відрізняється від клітинної оболонки еукаріотів. Її складають специфічні полімерні комплекси, яких немає в клітинних структурах інших організмів. Будова і хімічний склад оболонки є сталими для певних видів бактерій, на що зважають під час діагностування. Вважають, що саме оболонка визначає забарвлення бактерії за Грамом. Так називають розроблений данським мікробіологом X. Грамом у 1884 р. метод фарбування мікроорганізмів, який дає змогу диференціювати бактерії.
Після забарвлення бактерій карболовим генціанвіолетом і обробки препаратів розчином йоду та промивання їх спиртом, клітини одних бактерій знебарвлюються, а інші залишаються синьо-фіолетовими. Саме за цією ознакою бактерії поділяють на дві групи: грампозитивні забарвлюються у синьо-фіолетовий колір, грамнегативні знебарвлюються. Суть фарбування мікробів за Грамом досі ще остаточно не з'ясована. Вважають, що в основі цього методу лежать будова і особливості хімічного складу клітинних оболонок бактерій.
Характер побудови клітинної оболонки є важливішою ознакою, ніж саме фарбування за Грамом. Це дозволило Н. Гіббонсу і Р. Муррею ще в 1978 р. запропонувати грамнегативні еубактерії віднести до відділу грацілікутних, а грампозитивні — до відділу фірмікутних бактерій. До цих двох відділів прокаріотів належить переважна більшість бактерій [18].
До складу оболонки прокаріотів входять сім груп хімічних речовин (табл. 1). Як видно, хімічний склад оболонки грампозитивних і грамнегативних еубактерій досить помітно відрізняється. У перших основним компонентом клітинної стінки є пептидоглікани, які ще називають глікопептидами, мукопептидами, або муреїнами. У других — грамнегативних прокаріотів — вміст цього структурного полімеру в кілька разів менший, відсутні тейхоєві кислоти тощо.
Вважають, що головним структурним компонентом оболонок більшості бактерій є пептидоглікан (муреїн). Тільки у мікоплазм, L-форм бактерій, архебактерій та деяких метаноутворюючих і гало-фільних бактерій він відсутній. Пептидоглікан — гетерополімер, який складається з лінійних молекул глікану. Полісахаридний кістяк муреїну побудований із залишків, які чергуються, N-ацетилглюкозо-аміну і N-ацетилмурамової кислоти, з'єднаних між собою за допомогою β-1,4-глікозидних зв'язків (рис. 2). У грампозитивних бактерій нині виявлено близько 100 різних хімічних типів муреїну.
Таблиця 1Хімічний склад клітинних оболонок грампозитивних і грамнегативних еубактерій
| Компоненти клітинної оболонки | Грампозитивні еубактерії | Грамнегативні еубактерії | |
| Внутрішній пептидоглікановий) шар | Зовнішній шар (зовнішня клітинна мембрана) | ||
| Пептидоглікан | + | + | - |
| Тейхоєві кислоти | + | - | - |
| Полісахариди | + | - | + |
| Білки | ± | - | + |
| Ліпіди | ± | - | + |
| Ліпополісахариди | - | - | + |
| Ліпопротеїни | - | ± | + |
Примітка: + — наявні у малій кількості, ± — наявне в усіх видів, — відсутні.
До сітки пептидоглікану клітинної оболонки прокаріотів входять також тейхоєві і тейхуронові кислоти, поліпептиди, ліпополісахариди, ліпопротеїни та ін. Тейхоєві кислоти після пептидоглікану є другим унікальним класом хімічних сполук клітинної стінки грампозитивних бактерій. Це полімери, які побудовані на основі спиртів рибіту і гліцерину, що їхні залишки з'єднані між собою фосфодіефір-ними зв'язками. При цьому утворюються рибіттейхоєві та гліцерол-тейхоєві кислоти.
Крім тейхоєвих кислот, у оболонках грампозитивних бактерій виявлено тейхуронові кислоти, що утворюються із залишків уронових кислот та N-ацетилглюкозоаміну. В разі бракування фосфору у середовищі вони спроможні заміняти тейхоєві кислоти, які ковалентно можуть з'єднуватися з N-ацетилмурамовою кислотою, їхні довгі лінійні молекули інколи пронизують увесь пептидоглікановий шар, сягаючи зовнішньої поверхні оболонки. Вільні гідроксили фосфорної кислоти, які залишаються, надають тейхоєвим кислотам властивості поліаніонів, завдяки чому ці кислоти визначають поверхневий заряд клітини.

Рис. 2. Структура повторювальної одиниці пептидоглікану клітинної оболонки еубактерій.
Цифри в кільцях позначають: 1,2 — місця полімеризації гліканового скелета молекули; 3 - місце приєднання за допомогою фосфодіефірпого зв'язку молекули тейхоєвої кислоти в клітинній стінці грампозитивних бактерій; 4, 5 - місця, де відбуваються зв'язування між глікановими ланцюгами за допомогою пептидних зв'язків; 6 - місце ковалентного зв'язування з ліпонротеіном зовнішньої мембрани у грамнегативних бактерій; 7— місце дії лізоциму
У складі клітинної оболонки грампозитивних бактерій також виявлено в невеликих кількостях білки, ліпіди і полісахариди. З'ясовано, що полісахариди і ліпіди можуть ковалентно з'єднуватися з макромолекулами оболонки. Стосовно білків, то припускають, що вони виконують захисну функцію [18].
У грамнегативних еубактерій будова клітинної оболонки є набагато складнішою, ніж у грампозитивних (табл. 1). У середині оболонки цих бактерій міститься пептидоглікановий шар. Ззовні від нього розташований ще один шар (зовнішня мембрана), який складається з фосфоліпідів, ліпополісахаридів і білків, а під ним — цитоплазматична (внутрішня) мембрана, до складу якої також входять фосфоліпіди, білки тощо.
Білки зовнішньої мембрани грамнегативних бактерій поділяють на дві групи: основні, які беруть участь у формуванні мембранних гідрофільних пор (їх ще називають поринами) і мінорні білки, що виконують транспортну і рецепторну функції. Вони транспортують у клітину залізовмісні сполуки, вітаміни тощо.
Між зовнішньою і внутрішньою мембранами клітинної оболонки грамнегативних бактерій існує периплазматичний простір (пери-плазма), в якому, крім муреїнового шару, містяться специфічні білки, олігосахариди, неорганічні речовини і вода. Периплазматичні транспортні білки відіграють важливу роль у надходженні в клітину амінокислот, цукрів, фосфатів тощо.
Серед прокаріотів виявлено види бактерій, клітинна оболонка яких за структурою та хімічним складом помітно відрізняється від грампозитивних і грамнегативних типів. Вони належать до групи архебактерій, зокрема оболонки метаноутворюючих архебактерій містять пептидоглікан особливого хімічного складу. В інших представників цієї групи клітинна стінка складається переважно з кислого гетерополісахариду. Оболонка екстремальних галофілів, метаноутворюючих та ацидотермофільних архебактерій складається тільки з білка. Ці бактерії не забарвлюються за Грамом.
Поряд з цим слід зазначити, що за певних умов прокаріоти можуть існувати і без клітинних оболонок. Наприклад, за дії на клітини певними хімічними речовинами можна дістати структури (протопласти і сферопласти), які повністю або частково позбавлені оболонки. Вперше ці структури було виявлено у разі дії на бактерії ферментом лізоцимом з яєчного білка. Встановлено, що цей фермент розриває р-1,4-глікозидні зв'язки, які з'єднуют залишки N-ацетилглюкозоаміну і N-ацетилмурамової кислоти в пептидоглікані. Одержані при цьому протопласти або сферопласти набувають сферичної форми і в сприятливих умовах можуть виявляти певну метаболічну активність. Проте здатність до розмноження вони втрачають.
Унікальність структури і хімічного складу оболонки еубактерій та їх відмінність від рослинних і тваринних клітин дає змогу створювати і застосовувати медикаментозні препарати, які специфічно діють тільки на клітинну стінку прокаріотів і не завдають шкоди клітинам інших організмів. Прикладом цього є дія пеніциліну та деяких інших антибіотиків.
Цитоплазматична мембрана. Обов'язковим структурним компонентом бактеріальної клітини є цитоплазматична мембрана, яка відокремлює цитоплазму від клітинної оболонки. У мікоплазм та деяких інших прокаріотів цитоплазматична мембрана заміняє клітинну стінку. Під електронним мікроскопом цитоплазматична мембрана здебільшого має вигляд тришарового утворення з різною електронною щільністю, завтовшки 5—10 нм. На її частку припадає до 15 % сухої речовини клітини.
За хімічним складом цитоплазматична мембрана є білково-ліпідним комплексом, який містить 50-75 % білків, 15-45 % ліпідів та невеличку кількість вуглеводів. У ліпідах мембран багатьох прокаріотів виявлено низку специфічних жирних кислот, яких немає в мембранах еукаріотів. Це циклопропанові жирні кислоти. Головним ліпідним компонентом мембран бактерій є фосфоліпіди — похідні 3-фосфогліцеролу. Ліпіди підтримують механічну стабільність і зумовлюють гідрофобні властивості мембран [21].
На відміну від ліпідів, білковий склад цитоплазматичної мембрани прокаріотної клітини є набагато різноманітнішим. Наприклад, мембрана кишкової палички містить 27 основних і велику кількість мінорних білків. За амінокислотним складом ці білки майже не відрізняються від інших клітинних білків, за виключенням низького вмісту цистину в них. Досліди показали, що білки мембран бактеріальної клітини переважно є ферментами.
Донедавна найбільше визнання мала модель мембрани, запропонована ще 1935 р. Г.Доусоном і Д.Данієлі, яка дістала назву "елементарної мембрани". Згідно з цією моделлю, мембрана побудована з двох білкових шарів, між якими розташований бімолекулярний шар ліпідів. Однак останніми роками нагромадилось багато даних, які свідчать, що цитоплазматична мембрана має набагато складнішу будову, ніж "елементарна".
Тепер більшість вчених поділяють думку про те, що структура бактеріальної мембрани підпадає під рідинно-мозаїчну модель, розроблену для еукаріотів. Цитоплазматична мембрана утворена мінливим ліпідним бішаром, в який вмонтовані білки. Разом вони утворюють рухому мозаїку. Отже, за цією моделлю мембрана є "рідкою", лабільною, динамічною структурою, якій притаманна молекулярна асиметрія і мінливість.
При "біологічних" температурах мембранні ліпіди перебувають у розрідженому стані, який характеризується частковою впорядкованістю структури. Із зниженням температури вони переходять у кристалічний стан. "Рідка" структура мембран забезпечує свободу білкам, що є необхідним для здійснення процесів транспорту електронів і речовин через мембрану. Ця властивість також зумовлює високу еластичність мембран.
За сучаними даними, білки, що входять до складу цитоплазматичної мембрани, можна умовно поділити на такі групи: інтегральні, які цілком занурені в мембрану, а подекуди пронизують її наскрізь; периферійні білки, частково занурені в гідрофобну ділянку, і поверхневі, що містяться на поверхні мембрани (рис. 3). Зв'язок інтегральних білків з ліпідами частково, а периферійних повністю визначається електростатичними взаємодіями. Поряд з цим деякі білки і ліпіди н мембрані можуть бути ковалентно зв'язаними.
Отже, зовнішня цитоплазматична мембрана — складне високоорганізоване і високоспеціалізоване утворення, що виконує різноманітні функції. Вона є основним бар'єром, який забезпечує вибіркове надходження в клітину і вихід з неї різних речовин (іонів). На поверхні мембрани локалізовані ферментні системи, які беруть участь у синтезі мембранних ліпідів, компонентів клітинної оболонки та інших речовин. Клітинні мембрани містять високочутливі рецептори, за допомогою яких клітини розпізнають і, відповідним чином, реагують на інформацію, яка надходить ззовні [1].
Дуже важлива роль належить цитоплазматичній мембрані у перетворенні клітинної енергії. У бактерій, джерелом енергії для яких є процеси дихання або фотосинтезу, в цитоплазматичній мембрані пев ним чином розміщені переносники електронів (ЕТЛ), функціонування яких приводить до генерування електрохімічної енергії (AjlH+ ), що використовується потім у клітині по різних каналах, у тому числі й для утворення хімічної енергії (АТФ). У мембрані є також ферментні комплекси, які забезпечують перетворення AJIEP ї± АТФ.
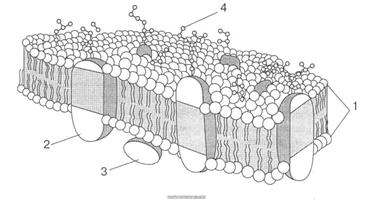
Рис. 3. Рідинно-мозаїчна структура мембран, запропонована С. Дж. Сінгером і Г. Ніколсоном (1972): 1 — біомолекулярний шар фосфоліпідів; 2 — внутрішній білок; 3 — периферійний білок; 4 — олігосахаридні групи
Цитоплазматична мембрана має спеціальні ділянки для прикріплення хромосоми і плазмід за реплікації їх і наступній сегрегації. Нарешті, мембрані належить інтегруюча роль в організмі, яка поєднується з бар'єрною функцією. Клітина являє собою єдине ціле, і порушення цілісності структури цитоплазматичної мембрани призводить до втрати нею життєдіяльності.
Внутрішньоцитоплазматичні мембрани. Розвинуті системи внутрішньоклітинних мембран мають фототрофні, хемотрофні та деякі інші бактерії. Виділяють кілька видів цих мембран. У фотосинтезуючих бактерій внутрішньоцитоплазматичні мембрани можуть мати форму трубочок, пласких дисків (тилакоїдів), пухирців (везикул, хроматофорів) тощо. Оскільки в цих мембранних структурах локалізовано фотосинтетичний апарат клітини (фотосинтезуючі пігменти та системи фосфорилювання), вони дістали загальну назву фотосинтетичних мембран. Всі фотосинтетичні мембрани є похідними цитоплазматичної мембрани, які виникли внаслідок її розростання та інвагінації в цитоплазму.
У зелених фототрофних бактерій та ціанобактерій внутрішньоклітинні фотосинтетичні мембрани відсутні. Основні компоненти фотосинтетичного апарату в них локалізовано на цитоплазматичній мембрані, а світловловлюючі пігменти містяться в органелах, які прилягають до мембрани — хлоросомах і фікобілісомах. У різних груп прокаріотів цитоплазматична мембрана утворює локальні впинання, які дістали назву мезосом. Ці мембранні утворення добре розвинуті у грампозитивних еубактерій. У грамнегативних вони трапляються рідше і менш розвинуті. За формою, розмірами і розміщенням розрізняють три основні типи мезосом: ламе-лярні (пластинчасті), тубулярні (трубчасті) і везикулярні, що нагадують пухирці. Є мезосоми й змішаного типу. Мезосоми найчастіше містяться в зоні формування клітинної перетинки і поділу бактеріального ядра.
Немає єдиної думки щодо функцій мезосом у прокаріотній клітині. Згідно з однією гіпотезою, мезосоми відіграють важливу роль у реплікації і сегрегації ДНК, поділі клітин і утворенні клітинної оболонки. За іншою гіпотезою, мезосоми не є обов'язковими структурами бактеріальної клітини. Вони тільки підсилюють певні клітинні функції. Нарешті, припускають, що мезосоми не беруть активної участі в процесах клітинного метаболізму, а виконують суто структурну функцію, що забезпечує компартменталізацію клітини, створює більш сприятливі умови для перебігу різних ферментативних реакцій [18].
Цитоплазма. Вміст клітини, оточений цитоплазматичною мембраною, називається цитоплазмою. За фізико-хімічною природою цитоплазма — складна колоїдна система. її складають: вода, білки, ліпіди, вуглеводи, мінеральні сполуки та інші речовини, співвідношення яких залежить від виду, віку, живлення бактерій та інших чинників. Фракція цитоплазми, яка має гомогенну консистенцію і містить макромолекули розчинних РНК, ферментних білків, продуктів і субстратів різних метаболічних реакцій тощо, дістала назву цитозоля.
Друга фракція цитоплазми містить різноманітні структурні утворення: внутрішньоцитоплазматичні мембрани, генетичний апарат (нуклеоїд і плазміди), рибосоми, інші внутрішньоклітинні структури і включення різної хімічної природи та функціонального призначення.
Рибосоми — округлі рибонуклепротеїнові тільця діаметром 15-20 нм, що розташовані довільно в цитоплазмі бактерій і складаються на 40 % із білка і на 60 % із РНК. їх кількість у клітині залежить від інтенсивності синтезу білка і може коливатися від 5000 до 90 000. Рибосоми прокаріотів мають константу седиментації 70 S , від чого і дістали назву 70 S-частинок. Вони побудовані з двох рибонуклеопротеї-нових субодиниць: малої 30 S і великої 50 S . Мала субодиниця побудована з однієї молекули 16 S РНК і переважно з однієї молекули кожного з білків 21 виду. Велика субодиниця містить дві молекули РНК (23 S і 5 S ) та по одній копії білків 34 видів. Більша частина рибосомальних білків виконує структурну функцію.
За коефіцієнтом седиментації* та деякими іншими властивостями, до бактеріальних рибосом подібні рибосоми мітохондрій і хлоропластів еукаріотних клітин. Рибосоми є центрами синтезу білка в клітині. Під час синтезу білка вони набувають форми агрегатів, які називаються полірибосомами. Ці полісоми містяться у цитоплазмі або зв'язані з мембранними структурами.
Генетичний апарат.
Тривалий час точилася дискусія про наявність у бактеріальній клітині ядра, подібного до ядра, що міститься в еукаріотних клітинах. Тепер ні в кого не виникає сумніву щодо наявності генетичного апарату в клітинах прокаріотів. Як і в еукаріотів він представлений молекулами ДНК, але між ними існують істотні відмінності у структурній організації. У прокаріотній клітині, на відміну від еукаріотної, ядерний апарат не відмежований від цитоплазми мембраною та займає певну ділянку в цитоплазмі. Враховуючи цю відмінність, ядерний апарат у прокаріотів запропоновано називати нуклеоїдом. Він міститься всередині цитоплазми і складається з гігантської молекули ДНК у формі замкненої в кільце подвійної спіралі, довжина якої у ціанобактерій може сягати 3 мм.
8-09-2015, 19:28