План.
Значение различных участков коры полушарий
головного мозга.
2. Двигательные функции.
3. Функции кожной и проприорицептивной
чувствительности.
4. Слуховые функции.
5. Зрительные функции.
6. Морфологические основы локализации функций в
коре головного мозга.
- Ядро двигательного анализатора
- Ядро слухового анализатора
- Ядро зрительного анализатора
- Ядро вкусового анализатора
- Ядро кожного анализатора
7. Биоэлектрическая активность головного мозга.
8. Литература.
ЗНАЧЕНИЕ РАЗЛИЧНЫХ УЧАСТКОВ КОРЫ БОЛЬШИХ
ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
С давних времен между учеными идет спор о местонахождении (локализации) участков коры головного мозга, связанных с различными функциями организма. Были высказаны самые разнообразные и взаимно противоположные точки зрения. Одни считали, что каждой функции нашего организма соответствует строго определенная точка в коре головного мозга, другие отрицали наличие каких бы то ни было центров; любую реакцию они приписывали всей коре, считая ее целиком однозначной в функциональном отношении. Метод условных рефлексов дал возможность И. П. Павлову выяснить ряд неясных вопросов и выработать современную точку зрения.
В коре головного мозга нет строго дробной локализации фун кций. Это следует из экспериментов над животными, когда после разрушения определенных участков коры, например двигательного анализатора, через несколько дней соседние участки берут на себя функцию разрушенного участка и движения животного восстанавливаются.
Эта способность корковых клеток замещать функцию выпавших участков связана с большой пластичностью коры головного мозга.
И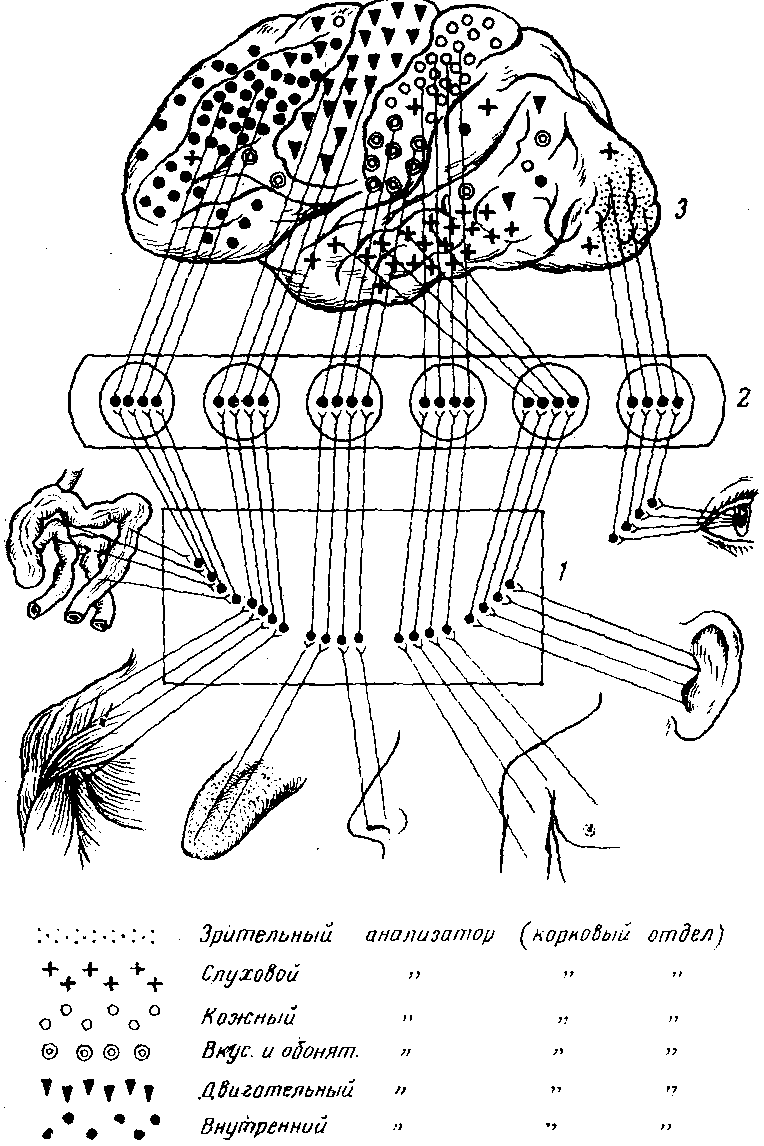 .
П. Павлов считал,
что отдельные
области коры
имеют разное
функциональное
значение. Однако
между этими
областями не
существует
строго определенных
границ. Клетки
одной области
переходят в
соседние области.
.
П. Павлов считал,
что отдельные
области коры
имеют разное
функциональное
значение. Однако
между этими
областями не
существует
строго определенных
границ. Клетки
одной области
переходят в
соседние области.
Рисунок 1. Схема связи отделов коры с рецепторами.
1 – спинной или продолговатый мозг; 2 – промежуточный мозг; 3 – кора головного мозга
В центре этих областей находятся скопления наиболее специализированных клеток-так называемые ядра анализатора, а на периферии-менее специализированные клетки.
В регуляции функций организма принимают участие не строго очерченные какие-то пункты, а многие нервные элементы коры.
Анализ и синтез поступающих импульсов и формирование ответной реакции на них осуществляются значительно большими областями коры.
Рассмотрим некоторые области, имеющие преимущественно то или иное значение. Схематическое расположение местонахождения этих областей приведено на рисунке 1.
Двигательные функции. Корковый отдел двигательного анализатора расположен главным образом в передней центральной извилине, кпереди от центральной (роландовой) борозды. В этой области находятся нервные клетки, с деятельностью которых связаны все движения организма.
 Отростки
крупных нервных
клеток, находящихся
в глубоких
слоях коры,
спускаются
в продолговатый
мозг, где значительная
часть их перекрещивается,
т. е. переходит
на противоположную
сторону. После
перехода они
опускаются
по спинному
мозгу, где
перекрещивается
остальная
часть. В передних
рогах спинного
мозга они вступают
в контакт с
находящимися
здесь двигательными
нервными клетками.
Таким образом,
возбуждение,
возникшее в
коре, доходит
до двигательных
нейронов передних
рогов спинного
мозга и затем
уже по их волокнам
поступает к
мышцам. Ввиду
того что в
продолговатом,
а частично и
в спинном мозгу
происходит
переход (перекрест)
двигательных
путей на противоположную
сторону, возбуждение,
возникшее в
левом полушарии
головного
мозга, поступает
в правую половину
тела, а в левую
половину тела
поступают
импульсы из
правого полушария.
Вот почему
кровоизлияние,
ранение или
какое-либо
другое поражение
одной из сторон
больших полушарий
влечет за собой
нарушение
двигательной
деятельности
мышц противоположной
половины тела.
Отростки
крупных нервных
клеток, находящихся
в глубоких
слоях коры,
спускаются
в продолговатый
мозг, где значительная
часть их перекрещивается,
т. е. переходит
на противоположную
сторону. После
перехода они
опускаются
по спинному
мозгу, где
перекрещивается
остальная
часть. В передних
рогах спинного
мозга они вступают
в контакт с
находящимися
здесь двигательными
нервными клетками.
Таким образом,
возбуждение,
возникшее в
коре, доходит
до двигательных
нейронов передних
рогов спинного
мозга и затем
уже по их волокнам
поступает к
мышцам. Ввиду
того что в
продолговатом,
а частично и
в спинном мозгу
происходит
переход (перекрест)
двигательных
путей на противоположную
сторону, возбуждение,
возникшее в
левом полушарии
головного
мозга, поступает
в правую половину
тела, а в левую
половину тела
поступают
импульсы из
правого полушария.
Вот почему
кровоизлияние,
ранение или
какое-либо
другое поражение
одной из сторон
больших полушарий
влечет за собой
нарушение
двигательной
деятельности
мышц противоположной
половины тела.
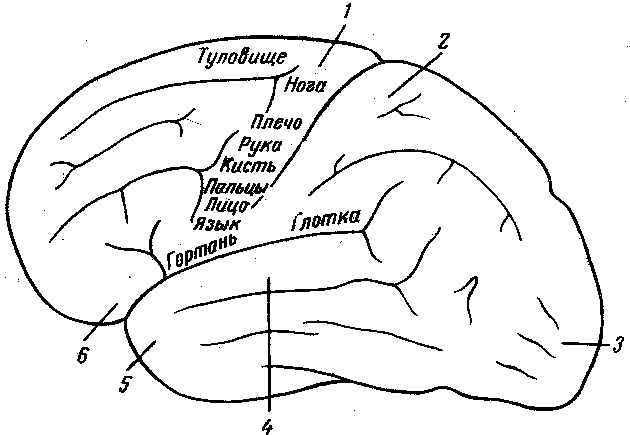
Рисунок 2. Схема отдельных областей коры больших полушарий головного мозга.
1 – двигательная область;
2 – область кожной
и проприорицептивной чувствительности;
3 – зрительная область;
4 – слуховая область;
5 – вкусовая область;
6 – обонятельная область
В передней центральной извилине центры, иннервирующие разные мышечные группы, расположены так, что в верхней части двигательной области находятся центры движений нижних конечностей, затем ниже-центр мышц туловища, еще ниже-центр передних конечностей и, наконец, ниже всех-центры мышц головы.
Центры разных мышечных групп представлены неодинаково и занимают неравномерные области.
Функции кожной и проприоцептивной чувствительности. Область кожной и проприоцептивной чувствительности у человека находится преимущественно позади центральной (роландовой) борозды в задней центральной извилине.
Локализация этой области у человека может быть установлена методом электрического раздражения коры головного мозга во время операций. Раздражение различных участков коры и одновременньш опрос больного об ощущениях, которые он при этом испытывает, дают возможность составить довольно четкое представление об указанной области. С этой же областью связано так называемое мышечное чувство. Импульсы, возникающие в проприорецепторах-рецепторах, находящихся в суставах, сухожилиях н мышцах, поступают преимущественно в этот отдел коры.
Правое полушарие воспринимает импульсы, идущие по центростремительным волокнам преимущественно с левой, а левое полушарие-преимущественно с правой половины тела. Этим объясняется то, что поражение, допустим, правого полушария вызовет нарушение чувствительности преимущественно левой стороны.
Слуховые функции. Слуховая область расположена в височной доле коры. При удалении височных долей нарушаются сложные звуковые восприятия, так как нарушается возможность анализа и синтеза звуковых восприятий.
Зрительные функции. Зрительная область находится в затылочной доле коры головного мозга. При удалении затылочных долей головного мозга у собаки наступает потеря зрения. Животное не видит, натыкается на предметы. Сохраняются только зрачковые рефлексы У человека нарушение зрительной области одного из полушарий вызывает выпадение половины зрения каждого глаза. Если поражение коснулось зрительной области левого полушария, то выпадают функции носовой части сетчатки одного глаза и височной части сетчатки другого глаза.
Такая особенность поражения зрения связана с тем, что зрительные нервы по пути к коре частично перекрещиваются.
Морфологические основы динамической локализации функций в коре полушарий большого мозга (центры мозговой коры).
Знание локализации функций в коре головного мозга имеет огромное теоретическое значение, так как дает представление о нервной регуляции всех процессов организма и приспособлении его к окружающей среде. Оно имеет и большое практическое значение для диагностики мест поражения в полушариях головного мозга.
Представление о локализации функций в коре головного мозга связано прежде всего с понятием о корковом центре. Еще в 1874 г. киевский анатом В. А, Бец выступил с утверждением, что каждый учасгок коры отличается по строению от других участков мозга. Этим было положено начало учению о разнокачественности коры головного мозга - цитоархитектонике (цитос - клетка, архитектонес - строю). В настоящее время удалось выявить более 50 различных участков коры - корковых цитоархитектонических полей, каждое из которых отличается от других по строению и расположению нервных элементов. Из этих полей, обозначаемых номерами, составлена специальная карта мозговой коры человека .
П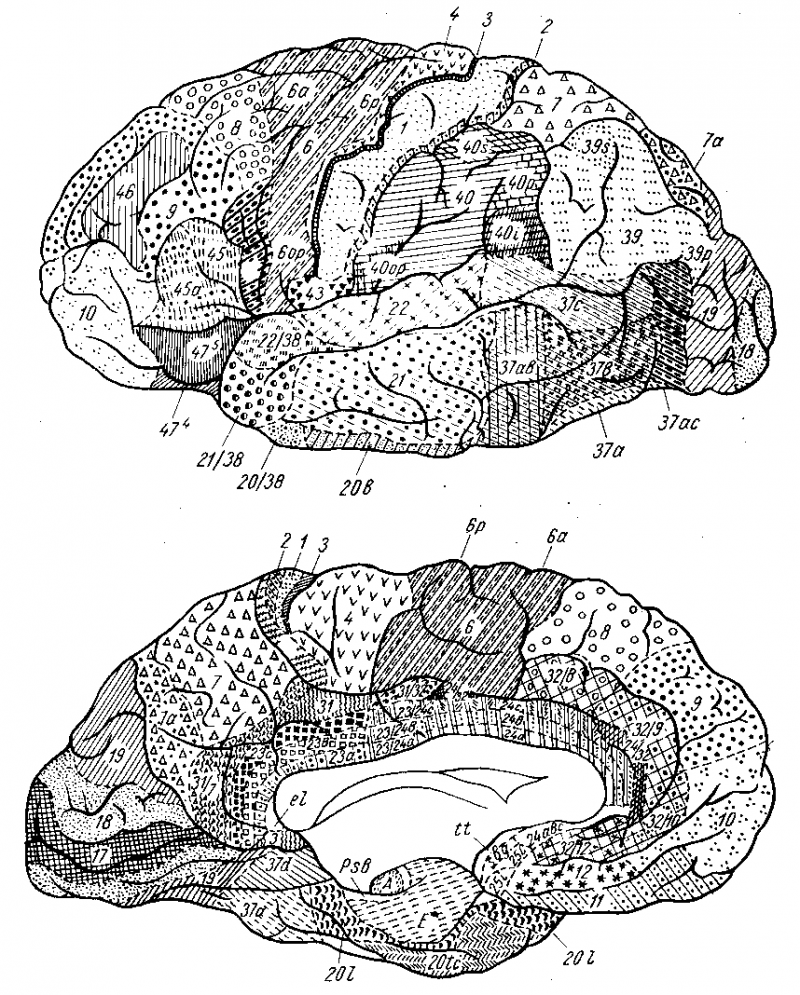 о
И.П.Павлову,
центр-это мозговой
конец так называемого
анализатора.
Анализатор
- это нервный
механизм, функция
которого состоит
в том, чтобы
разлагать
известную
сложность
внешнего и
внутреннего
мира на отдельные
элементы, т. е.
производить
анализ. Вместе
с тем благодаря
широким связям
с другими
анализаторами
здесь происходит
и синтезирование
анализаторов
друг с другом
и с разными
деятельностями
организма.
о
И.П.Павлову,
центр-это мозговой
конец так называемого
анализатора.
Анализатор
- это нервный
механизм, функция
которого состоит
в том, чтобы
разлагать
известную
сложность
внешнего и
внутреннего
мира на отдельные
элементы, т. е.
производить
анализ. Вместе
с тем благодаря
широким связям
с другими
анализаторами
здесь происходит
и синтезирование
анализаторов
друг с другом
и с разными
деятельностями
организма.
Рисунок 3. Карта цитоархитектонических полей мозга человека (по данным института моэга АМН СССР) Вверху - верхнелатеральная поверхность,внизу- медиальная поверхносгь. Объяснение в тексте.
В настоящее время вся мозговая кора рассматривается как сплошная воспринимающая поверхность. Кора - это совокупность корковых концов анализаторов. С этой точки зрения мы и рассмотрим топографию корковых отделов анализаторов, т. е. главнейшие воспринимающие участки коры полушарий большого мозга.
Прежде всего рассмотрим корковые концы анализаторов, воспринимающих раздражения из внутренней среды организма .
1. Ядро двигательного анализатора, т. е. анализатора проприоцептивных (кинестетических) раздражении, исходящих от костей, суставов, скелетных мышц и их сухожилий, находится в предцентральной извилине (поля 4 и 6} и lobulus paracentralis. Здесь замыкаются двигательные условные рефлексы. Двигательные параличи, возникающие при поражении двигательной зоны, И. П. Павлов объясняет не повреждением двигательных эфферентных нейронов, а нарушением ядра двигательного анализатора, вследствие чего кора не воспринимает кинестетические раздражения и движения становятся невозможными. Клетки ядра двигательного анализатора заложены в средних слоях коры моторной зоны. В глубоких ее слоях (V, отчасти VI) лежат гигантские пирамидные клетки, представляющие собой эфферентные нейроны, которые И. П. Павлов рассматривает как вставочные нейроны, связывающие кору мозга с подкорковыми ядрами, ядрами черепных нервов и передними рогами спинного мозга, т. е. с двигательными нейронами. В предцентральной извилине тело человека, так же как и в задней, спроецировано вниз головой. При этом правая двигательная область связана с левой половиной тела и наоборот, ибо начинающиеся от нее пирамидные пути перекрещиваются частью в продолговатом, а частью в спинном мозге. Мышцы туловища, гортани, глотки находятся под влиянием обоих полушарий. Кроме предцентральной извилины, проприоцептивные импульсы (мышечно-суставная чувствительность) приходят и в кору постцентральной извилины.
2. Ядро двигательного анализатора, имеющего-отношение к сочетанному повороту головы и глаз в противоположную сторону, помещается в средней лобной извилине, в премоторной области (поле 8). Такой поворот происходит и при раздражении поля 17, расположенного в затылочной доле в соседстве с ядром зрительного анализатора. Так как при сокращении мышц глаза в кору мозга (двигательный анализатор, поле 8) всегда поступают не только импульсы от рецепторов этих мышц, но и импульсы от еет-чатки (зрительный анализатор, поле 77), то различные зрительные раздражения всегда сочетаются с различным положением глаз, устанавливаемым сокращением мышц глазного яблока.
3. Ядро двигательного анализатора, посредством которого происходит синтез целенаправленных сложных профессиональных, трудовых и спортивных движений, помещается в левой (у правшей) нижней теменной дольке, в gyrus supramarginalis (глубокие слои поля 40). Эти координированные движения, образованные по принципу временных связей и выработанные практикой индивидуальной жизни, осуществляются через связь gyrus supramarginalis с предцентральной извилиной. При поражении поля 40 сохраняется способность к движению вообще, но появляется неспособность совершать целенаправленные движения, действовать - апраксия (праксия - действие, практика).
4. Ядро анализатора положения и движения головы - статический анализатор (вестибулярный аппарат) в коре мозга точно еще не локализован. Есть основания предполагать, что вестибулярный аппарат проецируется в той же области коры, что и улитка, т. е. в височной доле. Так, при поражении полей 21 и 20, лежащих в области средней и нижней височных извилин, наблюдается атаксия, т. е расстройство равновесия, покачивание тела при стоянии. Этот анализатор, играющий решающую роль в прямохождении человека, имеет особенное значение для работы летчиков в условиях реактивной авиации, так как чувствительность вестибулярного аппарата на самолете значительно понижается.
5. Ядро анализатора импульсов, идущих от внутренностей и сосудов, находится в нижних отделах передней и задней центральных извилин. Центростремительные импульсы от внутренностей, сосудов, непроизвольной мускулатуры и желез кожи поступают в этот отдел коры, откуда отходят центробежные пути к подкорковым вегетативным центрам.
В премоторной области (поля 6 и 8) совершается объединение вегетативных функций.
Нервные импульсы из внешней среды организма поступают в корковые концы анализаторов внешнего мира.
1. Ядро слухового анализатора лежит в средней части верхней височной извилины, на поверхности, обращенной к островку, - поля 41, 42, 52, где проецирована улитка. Повреждение ведет к глухоте.
2. Ядро зрительного анализатора находится в затылочной доле - поля 18, 19. На внутренней поверхности затылочной доли, по краям sulcus Icarmus, в поле 77 заканчивается зрительный путь. Здесь спроецирована сетчатка глаза. При поражении ядра зрительного анализатора наступает слепотa. Выше поля 17 расположено поле 18, при поражении которого зрение сохраняется и только теряется зрительная память. Еще выше находится поле при поражении которого утрачивается ориентация в непривычной обстанвке.
3. Ядро вкусового анализатора, по одним данным, находится в нижней постцентральной извилине, близко к центрам мышц рта и языка, по другим - в ближайшем соседстве с корковым концом обонятельного анализатора, чем объясняется тесная связь обонятельных и вкусовых ощу-ний. Установлено, что расстройство вкуса наступает при поражении поля 43.
Анализаторы обоняния, вкуса и слуха каждого полушария связаны с рецепторами соответствующих органов обеих сторон тела.
4. Ядро кожного анализатора (осязательная, болевая и температурная чувствительность) находится в постцентральной извилине (поля 7, 2, 3) и в пе верхней теменной области (поля 5 и 7).
Частный вид кожной чувствительности - узнавание предметов на ощупь - стереогнозия (стереос - пространственный, гнозис - знание) связана с участком коры верхней теменной дольки (поле 7) перекрестно: левое полушарие соответствует правой руке, правое - левой руке. При поражении поверхностных слоев поля 7 утрачивается способность узнавать предметы на ощупь, при закрытых глазах.
Биоэлектрическая активность головного мозга.
Отведение биопотенциалов головного мозга - электроэнцефалография-дает представление об уровне физиологической активности головного мозга. Кроме метода электроэнцефалографии-записи биоэлектрических потенциалов, используется метод энцефалоскопии-регистрации колебаний яркости свечения множества точек мозга (от 50 до 200).
Электроэнцефалограмма является интегративным пространственно-временным показателем спонтанной электрической активности мозга. В ней различают амплитуду (размах) колебаний в микровольтах и частоту колебаний в герцах. В соответствии с этим в электроэнцефалограмме различают четыре типа волн: -, -, - и -ритмы. Для -ритма характерны частоты в диапазоне 8-15 Гц, при амплитуде колебаний 50-100 мкВ. Он регистрируется только у людей и высших обезьян в состоянии бодрствования, при закрытых глазах и при отсутствии внешних раздражителей. Зрительные раздражители тормозят -ритм.
У отдельных людей, обладающих живым зрительным воображением, -ритм может вообще отсутствовать.
Для деятельного мозга характерен (-ритм. Это электрические волны с амплитудой от 5 до 30 мкВ и частотой от 15 до 100 Гц Он хорошо регистрируется в лобных и центральных областях головного мозга. Во время сна появляется -ритм. Он же наблюдается при отрицательных эмоциях, болезненных состояниях. Частота потенциалов -ритма от 4 до 8 Гц, амплитуда от 100 до 150 мкВ Во время сна появляется и -ритм - медленные (с частотой 0,5-3,5 Гц), высокоамплитудные (до 300 мкВ) колебания электрической активности мозга .
Помимо рассмотренных видов электрической активности, у человека регистрируется Е-волна (волна ожидания раздражителя) и веретенообразные ритмы. Волна ожидания регистрируется при выполнении сознательных, ожидаемых действий. Она предшествует появлению ожидаемого раздражителя во всех случаях, даже при неоднократном его повторении. По-видимому, ее можно рассматривать как электроэнцефалографический коррелят акцептора действия, обеспечивающего предвидение результатов действия до его завершения. Субъективная готовность отвечать на действие стимула строго определенным образом достигается психологической установкой (Д. Н. Узнадзе). Веретенообразные ритмы непостоянной амплитуды, с частотой от 14 до 22 Гц, появляются во время сна. Различные формы жизне деятельности приводят к существенному изменению ритмов биоэлектрической активности мозга.
При умственной работе усиливается -ритм, -ритм при этом исчезает. При мышечной работе статического характера наблюдается десинхронизация электрической активности мозга. Появляются быстрые колебания с низкой амплитудой.Во время динамической работы пе-. риоды десинхронизированной и синхронизированной активности наблюдаются соответственно в моменты рабогы и отдыха.
Образование условного рефлекса сопровождается десинхронизацией волновой активности мозга.
Десинхронизация волн происходит при переходе от сна к бодрствованию . При этом веретенообразные ритмы сна сменяются
-ритмом, увеличивается электрическая активность ретикулярной формации. Синхронизация (одинаковые по фазе и направлению волны)
характерна для тормозного процесса. Она наиболее отчетливо выражена при выключении ретикулярной формации стволовой части мозга. Волны электроэнцефалограммы, по мнению большинства исследователей, являются результатом суммации тормозных и возбуждающих постсинаптических потенциалов. Электрическая активность мозга не является простым отражением обменных процессов в нервной ткани. Установлено, в частности, что в импульсной активности отдельных скоплений нервных клеток обнаруживаются признаки акустического и семантического кодов.
Литература.
1. М.Г.Привес, Н.К.Лысенков, В.И.Бушкович. «Анатомия
человека».
2. Н.А.Фомин «Физоология человека»
3. А.А.Маркосян «Физиология».
Министерство высшего и профессионального образования
Российской Федерации
Амурский государственный университет
Кафедра МСР
Р е ф е р а т
По предмету: Физиология центральной нервной системы
На тему: Локализация функций в коре больших полушарий.
Электрическая активность головного мозга.
Выполнила: студентка 966 группы
Курицкая Галина Александровна
Проверил: преподаватель Кириченко Владимир Иванович
г. Благовещенск
2000 г.
8-09-2015, 21:01