Станкевич Л.И.
Клетки лимфоидной системы: взаимодействие во время развития иммунного ответа; пути физиологической регуляции.
Лимфоциты, как и все остальные элементы крови, образуются из популяции изначальных гематолимфоидных предшественников, так называемой стволовой кроветворной клетки (СКК). Дифференцировка СКК в Т- и В-клетки лимфоидного ряда начинается после ее попадания в особое индуцирующее дифференцировку микроокружение, имеющееся в таких органах, как костный мозг и тимус. Гипотетическая схема дифференцировки СКК и образования различных подклассов Т-лимфоцитов представлена на рис.1. Полипотентная СКК характеризуется тем, что одна из дочерних клеток, образовавшихся в результате ее деления, дифференцируется, а другая остается недифференцированной и служит источником образования последующих поколений дифференцированных клеток. СКК в результате такого деления может превратиться в самообновляющуюся клетку-предшественник с более ограниченными возможностями. Так, коммитированная стволовая лимфоцитарная клетка (СКЛ) дает начало пре-Т- и пре-В-клеткам. Более дифференцированная миелоидная стволовая клетка (СКМ) служит непосредственным предшественником других клеток крови (эритроциты, гранулоциты, мегакариоциты, тромбоциты и моноциты/макрофаги).
Схема дифференцировки лимфоидных клеток.
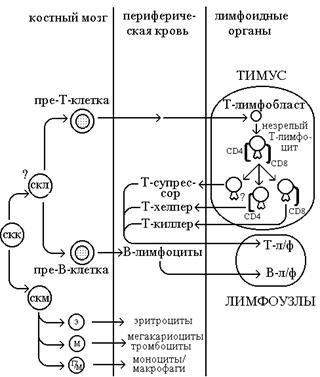
Обозначения:
СКК - стволовая кроветворная клетка
СКЛ - стволовая лимфоцитарная клетка
СКМ - стволовая миелоидная клетка
Предшественники Т-клеток, обнаруживаемые в гематопоэтической ткани, поступают в тимус, претерпевают там дифференцировку и выходят из тимуса уже в виде клеток с различными функциями, несущих на себе характерные маркеры. В тимусе происходит созревание и(или) отбор антиген-специфичных Т-клеток, а также избирательное высвобождение этих клеток на периферию.
Т-клеточные элементы иммунной системы являются ключевым звеном развития иммунологического ответа организма. Зрелые Т-лимфоциты разделяются на ряд подклассов в зависимости от иммунологических маркеров и выполняемых функций. Часть из них опосредует важные регуляторные реакции, в частности может "помогать" (хелперы) или "подавлять" (супрессоры) развитие иммунного ответа, в том числе образование антител В-лимфоцитами. Другие Т-лимфоциты выполняют эффекторные функции, например, вырабатывают растворимые вещества, запускающие разнообразные воспалительные реакции, или осуществляют прямое разрушение клеток, несущих на себе антигены ("киллерная" функция). В соответствии с этим различают Т-хелперы (несут Lyt1 маркер), Т-супрессоры и Т-киллеры (несут Lyt2 маркер), а также Т-клетки, участвующие в реакции замедленной гиперчувствительности и связанных с нею иммунологических явлениях .
Т-киллеры - наиболее агрессивные клетки этого звена. Они выискивают в других клетках признаки вирусной инфекции и иных аномалий (злокачественная трансформация). При встрече предшественника Т-киллера с антигенным комплексом (антиген, связанный с молекулой главного комплекса гистосовместимости (МНС)) на поверхности зараженной клетки прекиллер претерпевает многократное деление. Дочерние клетки несут рецепторы для этого антигена, становятся активными киллерами и разрушают клетки, зараженные этим вирусом, тем самым делая его уязвимым для антител, связывающих вирусные частицы (Фон Бёмер Х., Кисилов П., 1991). Распознавание антигена Т-киллерами специфично в отношении и антигенного пептида и молекул МНС. Специфичность, заключающаяся в двойном распознавании, обусловлена Т-клеточными рецепторами (TCR). Для полной активации прекиллера недостаточно связывания ТСR с антигенным комплексом. Необходимо связывание еще одной молекулы - CD8-корецептора. Этот белок одинаков у всех киллеров, но связывается в участке, отличном от TCR. СD8 - маркер Т-киллеров. Этот белок принимает участие в распознавании антигена киллерными клетками. Активация Т-киллера наиболее эффективна, когда TCR и CD8 связываются с одной и той же молекулой. Т-хелперные клетки тоже имеют TCR и инвариантный корецептор, которые совместно обеспечивают активацию Т-хелперов. Но в данном случае корецептор другой - CD4 (рис.2). ТСR и CD8 киллерных клеток связываются с молекулой МНС I класса и пептидами из белков, синтезирующихся в самой клетке. TCRи CD4 хелперов связываются с белками МНС II класса и антигенными пептидами из белков, поглощенных клеткой (DaynesR.A. etal., 1995). Т-хелперы отличаются по своей роли в иммунном ответе: они кооперируют с В-клетками в антительном ответе на такие антигены, как бактериальные токсины. Токсичный белок, попадающий в кровь, захватывается макрофагом. В-лимфоциты тоже поглощают молекулы токсинов, связавшиеся со специфичными рецепторами типа антител на их поверхности. Внутри макрофагов и В-лимфоцитов токсин расщепляется, а образующиеся пептиды презентируются на поверхность клетки белками МНС II класса. Как только ТСR и СD4 свяжутся с одной и той же молекулой МНС класса II на поверхности клетки, Т-лимфоциты начинают делиться, образуя активные хелперы (макрофаги максимально способны активировать эти клетки). Активные Т-хелперы секретируют интерлейкины (IL), которые стимулируют В-лимфоциты к делению и секреции больших количеств антител. Свободные антитела связывают токсин и нейтрализуют его.
Схема строения рецепторного аппарата Т-лимфоцитов.
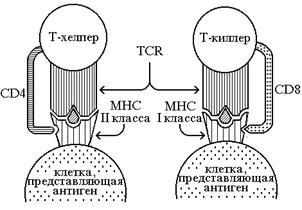
Обозначения:
![]() - антиген
- антиген
TCR - Т-клеточный рецептор
МНС - молекула основного
комплекса гистосовместимости
CD4, CD8 - маркерные корецепторы
Т-лимфоцитов
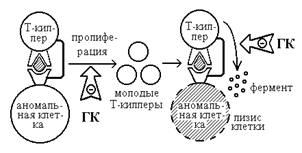
Основным физиологическим регулятором взаимодействия клеток во время формирования иммцнного ответа являются гормоны надпочечников - глюкокортикоиды (ГК). Ранние этапы развития иммунологических реакций, важнейшее звено которых - пролиферация клеток, являются наиболее чувствительными к действию ГК (рис.3). Важной стороной действия фармакологических доз ГК гормонов, во многом определяющей их тормозящее влияние на гуморальный и клеточный иммунный ответ, является способность гормонов угнетать процессы деления иммуннокомпетентных клеток. Установлено, что наиболее чувствительными к действию гормонов являются начальные фазы процесса пролиферации. Эффекты действия ГК на процессы деления клеток во многом зависят от способности этих гормонов подавлять продукцию интерлейкина-1 и интерлейкина-2, играющих ключевую роль в реализации пролиферативных реакций. Кроме интерлейкинов, ГК гормоны ингибируют продукцию и других гуморальных факторов, вырабатываемых активированными клетками иммунной системы.
Цитотоксические Т-лимфоциты также играют важнейшую роль в реализации клеточных иммунных реакций. Различные субпопуляции этих клеток осуществляют антителозависимый, не требующий присутствия комплимента лизис клеток-мишеней, то есть разрушение аутологичных, аллогенных или ксеногенных клеток в присутствии специфических против этих клеток антител или независимый от антител и комплимента лизис любых опухолевых клеток-мишеней аллогенной, ксеногенной или аутологической природы. ГК способны существенно тормозить функции всех субпопуляций этих лимфоцитов. Cуммарная схема ингибирования глюкокортикоидами передачи иммунного сигнала между лимфоидными клетками представлена на рис.3.
Пути ингибирования глюкокортикоидами передачи иммунного сигнала между лимфоидными клетками.
![]()
8-09-2015, 21:31