периферической крови
М.З.Федорова, В.Н.Левин, В.Д.Горичева, О.Н.Семенова
Согласно современным представлениям сезонные ритмы являются одной из форм приспособительных реакций организма к циклическим изменениям окружающей среды и присущи всем уровням биологической организации. Циркануальные колебания выявлены и описаны для многих показателей системы крови и иммунитета. Вместе с тем, в хронобиологических исследованиях практически отсутствуют сведения о сезонных изменениях неспецифической функциональной активности лейкоцитов, в частности, взаимосвязанных стадий фагоцитарного процесса (движение, адгезия, поглотительная способность), и сопротивляемости воздействиям окружающей среды. Целью проведенной работы было изучение физиологических реакций лейкоцитов крови интактных белых крыс в зимний (январь-февраль) и летний (май-июнь) периода.
Материал и методы исследования
Опыты проведены на 24 беспородных крысах-самцах 300-400 г. Кровь для исследований брали путем декапитации у предварительно наркотизированных животных. В качестве антикоагулянта использовали гепарин (10 ед./мл). Кровь центрифугитровали, собирали лейкоциты. Примесь эритроцитов разрушали 0,83% раствором хлорида аммония. Клетки дважды отмывали изотоничным буфером (раствор Дюльбекко). Отмытые клетки ресуспензировали в этом же растворе.
Для изучения локомоционной активности использовали метод миграции под агарозой [1]. В работе применяли два стекла. На первом в агаровом геле вырезали одиночную лунку, куда помещали суспензию лейкоцитов (7-10 х 105 клеток). По площади распространения клеток оценивали спонтанную миграцию. Для оценки индуцированной миграции на втором стекле вырезали группу из двух лунок – одну для клеточной суспензии, вторую – для хемоаттрактанта. В качестве последнего использовали свежую аутоплазму.
Адгезионную способность оценивали по методу, описанному [2]. Подсчитывали число клеток в исходной суспензии. После инкубации в термостате, капилляр с лейкоцитами перфузировали раствором Дюльбекко при напряжении сдвига 30 Н/м2. Определяли число клеток в смыве. Рассчитывали количество лейкоцитов, оставшихся в капилляре (клетки с большой силой сцепления).
Для изучения поглотительной способности использовали частицы латекса (0,8 мкм) [3]. Определяли число (%) фагоцитирующих (фагоцитарная активность) и среднее число поглощенных частиц (фагоцитарный индекс).
Определение осмотической стойкости лейкоцитов проводили классическим способом по методу Storti [4]. Процент клеток, сохранившихся после часовой инкубации в 0,2% растворе хлорида натрия служил показателем осмотической резистентности.
Ореолообразующую способность оценивали по способу, описанному в [5]. Процент ореолообразующих клеток через 10 мин. инкубации характеризовал скорость ореолообразования, а через 2-24 часа – ореолообразующую активность.
Морфометрические характеристики и механические свойства оценивали в опытах при перемещении лейкоцитов в стеклянных капиллярах с сужением (4 мкм) при постоянном отрицательном давлении 60 мм вод. ст. [6]. Определяли объем и площадь поверхности покоящихся (сферических) и деформированных (эллипсоидных) клеток, а также время восстановления до исходной формы.
Все полученные результаты обработаны статистически. Достоверность различий определяли по критерию t Стьюдента. Наличие связей между отдельными признаками оценивали по коэффициенту линейной корреляции.
Результаты исследований и их обсуждение
Сравнительный анализ данных, полученных при изучении морфометрических характеристик, функциональных свойств и реактивности лейкоцитов интактных лабораторных крыс в зимний и летний периоды показал наличие сезонных колебаний неспецифических физиологических реакций белых клеток крови. В летние месяцы установлено существенное снижение общей локомоционной а фагоцитарной активности полиморфно-ядерных лейкоцитов (рис. 1 и 2): площадь распространения клеток при спонтанной и стимулированной миграции в зимний период составляла 6,7±0,6 мм2 и 7,7±0,5 мм2, летом – 3,5±0,2 мм2 и 3,7±0,2 мм2; число фагоцитирующих нейтрофилов, соответственно - 84±1,6% и 75±1,7%.

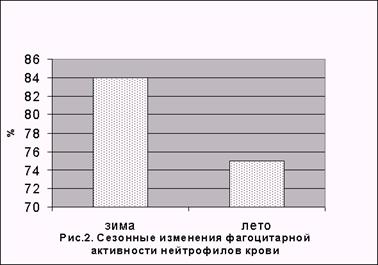
Меньшими, по сравнению с зимним периодом, были также показатели реактивности белых клеток крови. Зарегистрировано значительное снижение осмотической стойкости и ореолообразования (табл. 1).
Таблица 1
Реакции лейкоцитов в средах с низкой и высокой осмолярностью
| Сезон года | Осмотичкеская резистентность (%) | Скорость ореолообразования (%) | Ореолообразующая активность (%) |
| Зима | 70±7 | 0,6±0,05 | 6,2±1,90 |
| Лето | 47±5** | 0,3±0,05** | 1,0±0,06** |
Примечание: звездочка – достоверность различий по сравнению с зимним периодом (р<0,01).
Относительно стабильными в течение года оставались морфометрические параметры клеток в естественном “пассивном” и деформированном состоянии (объем и площадь поверхности), скорость восстановления исходной формы лейкоцитов после деформации, адгезионная способность. Незначительные сезонные колебания размера лейкоцитов проявлялись как тенденция (табл. 2).
Таблица 2
Показатели морфометрических характеристик и функциональных свойств лейкоцитов крови, имеющие относительно стабильные величины в разные сезоны года
| Сезон года | Объем лейкоцитов (мкм3) | Площадь поверхности клеток (мкм2) | Изменения площади поверхности при деформации (%) | Время восстановления исходной формы после деформации (с) | Доля адгезирующих клеток (%) |
| Зима | 189±26 | 152±14 | 31±5,6 | 59±8,2 | 51±8,6 |
| Лето | 170±19 | 147±17 | 21±4,4 | 63±8,1 | 51±4,3 |
Единственным функциональным показателем, величина которого в летний период оказалась выше, чем зимой, был фагоцитарный индекс (7,5±0,4 против 6,3±0,1, р<0,01, рис.3).
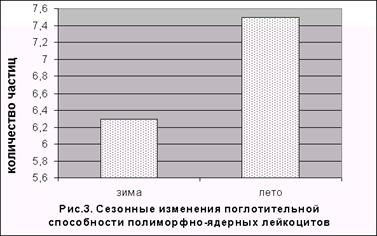
Корреляционный анализ результатов исследования позволил выявить различия статистических взаимосвязей между изученными показателями в разные сезоны года. Для зимнего периода характерино наличие сильных достоверных (р<0,05) связей между параметрами, отражающими функциональную активность лейкоцитов (табл.3). Летом они не зарегистрированы. Такого рода различия свидетельствуют о значительной напряженности в зимние месяцы физиологических реакций клеток, выполняющих в организме защитные функции. Результаты, демонстрирующие усиление защитных свойств организма в зимний период описаны и другими авторами. Установлено увеличение доли Т- и В-клеток осенью и зимой за счет сокращения количества малодифференцированных лимфоцитов (0-клеток) [7], а также числа нейтрофилов [8]. [9] отмечено не только накопление Т- и В-лимфоцитов, но их активизация с повышением уровня иммуноглобулинов G и М.
Таблица 3
Коррелирующие показатели функциональных свойств лейкоцитов крови в зимний период
| Показатели | Значения коэффициента корреляции | |
| 1 | 2 | |
| Ореолообразующая активность | Осмотическая резистентность | 0,89 |
| Время восстановления после деформации | 0,90 | |
| Время восстановления после деформации | Фагоцитарная активность | -0,86 |
| Площадь поверхности клеток в исходном состоянии | 0,78 | |
| Площадь поверхности деформированных клеток | 0,77 | |
| Площадь растпространения клеток при спонтанной миграции | Число адгезировавших клеток | -0,79 |
В исследовании [10] в зимнее время зарегистрирован наибольший титр комплемента и лизоцима.
Полученные в исследовании данные позволяют констатировать, что усиление неспецифической функциональной активности гранулоцитов и реактивности полиморфно-ядерных лейкоцитов и лимфоцитов в зимние месяцы происходит на фоне напряжения адаптационных реакций, вызванного сезонными изменениями метеорологических параметров.
Список литературы
Безносенко С.А., Барсуков А.А., Земсков В.М. Хемотаксис фагоцитов под агарозой. Методические аспекты//Журн. микробиологии. Эпидемиологии и иммунобиологии. 1984. №6. С. 101-105.
Mege J.-L.,Eon B., Saux P et al. Inhibition of Granulocyte Adhesion by Pentoxifylline//Proccedings of the Workshop on Pentoxifylline and Analogues: Effect on Leukocyte Function, 10-11 November 1989. Saint Paul-de-Vence, France. P. 17-23.
Потапова С.Г., Хрустиков В.С., Демидова Н.В., Козинец Г.И. Изучение поглотительной способности нейтрофилов крови с использованием инертных частиц латекса// Проблемы гематологии и переливания крови. 1977. №9. С. 58-59.
Storti E, Pederzini A. Augmentation de la resistance des globules blancs par l’administration de stiblene// Schweiz. Med. Wochenschr. 1955. V. 85. P. 38-39.
Кашулина А.П., Александрова Л.М.,Терещенко И.П. и др. Функциональное состояние нейтрофильных гранулоцитов периферической крови, определяемое по показателю ореолообразования, в процессе опухолевого роста// Патол. физиол. и эксперим. терапия. 1990. № 4. С. 37-39.
Tran-Son-Tay R., Needham D., Yeung A., Hochmuth R.M. Time-dependent recovery of passive neutrophils after large deformation// Biophys. J. 1991. V. 60. P. 856-866.
Деряпа Н.Р., Мошкин М.П., Посный В.С. Проблемы медицинской биоритмологии. М.: Медицина, 1985. 208 с.
Физиология лейкоцитов человека. Л.: Наука, 1979. 232 с.
Земсков В.М., Земсков Л.М., Золоедов В.И. Особенности иммунного реагирования при физиологических и патологических процессах// Экология человека. 1997. № 2. С. 37-41.
Кузьмин П.Н. Сезонные биоритмы иммунобиологической реактивности организма в условиях Западной Сибири// Климато-медицинские проблемы и вопросы медицинской географии Сибири. Томск, 1974, т. 1. С. 120-122.
8-09-2015, 21:43