Днепропетровская государственная медицинская академия
Кафедра анестезиологии и интенсивной терапии
Зав. Каф: проф., д.м.н. Л.В. Усенко
РЕФЕРАТ
На тему:
“Особенности проведения анестезиологического пособия в нейрохирургии ”
Кировоград - 2008
Введение
Для анестезиологического обеспечения нейрохирургических операций необходимо знание физиологии ЦНС. Многие анестетики оказывают значительное воздействие (как нежелательное, так и благоприятное) на метаболизм мозга, мозговой кровоток, образование и всасывание цереброспинальной жидкости, объем внутричерепного содержимого и внутричерепное давление. В настоящей главе обсуждаются важные для анестезиологической практики нейрофизиологические концепции, а также влияние анестетиков и различных вспомогательных средств на ЦНС. И хотя речь пойдет в основном о головном мозге, почти все изложенное можно отнести и к спинному мозгу.
ФИЗИОЛОГИЯ ЦНС
Метаболизм мозга
В покое мозг потребляет до 20 % получаемого организмом человека кислорода. Главный потребитель энергии в мозге — это фермент АТФ-аза, поддерживающий электрическую активность нейронов. Метаболизм в мозге обычно оценивают по потреблению кислорода мозгом, или CMRO2 (CMR — cerebral metabolic rate), которое у взрослых составляет в среднем 3-3,5 мл/100 г/мин (50 мл/мин). CMRO2 максимально в сером веществе коры больших полушарий и прямо пропорционально биоэлектрической активности коры. Потребность мозга в кислороде по сравнению с таковой у других органов чрезвычайно велика, а запасов кислорода в нем нет. Если кровь не поступает в мозг в течение 10 с, напряжение кислорода падает ниже 30 мм рт. ст. и человек теряет сознание. Если мозговой кровоток не восстанавливается в течение 3-8 мин, то запасы АТФ истощаются и возникает необратимое повреждение нейронов. Наиболее чувствительны к гипоксии нейроны гиппокампа pi мозжечка.
Нейроны обеспечиваются энергией преимущественно за счет утилизации глюкозы. В среднем по- требление глюкозы мозгом равно 5 мг/100 г/мин. В ЕЩС почти вся глюкоза (> 90 % ) подвергается аэробному распаду, поэтому потребление кислорода и потребление глюкозы изменяются параллельно. При голодании главным источником энергии для мозга становятся кетоновые тела — ацетоаце-тат и |3-гидроксибутират. Хотя мозг способен поглощать из крови молочную кислоту, ее метаболизм не играет значительной роли в энергетическом обеспечении. Не меньше чем гипоксия, опасна для мозга острая устойчивая гипогликемия. Исследования выявили парадоксальное явление: при тотальной ишемии мозга гипергликемия способствует внутриклеточному ацидозу и усугубляет повреждение нейронов. Влияние гипергликемии на очаговую ишемию не исследовано.
МОЗГОВОЙ КРОВОТОК
Мозговой кровоток (MK) зависит от интенсивности метаболизма. Мозговой кровоток чаще всего изучают с помощью изотопных методов исследований (обычно измеряют у~излучение изотопа ксенона [153 Xe]). После в/в инъекции изотопа датчики, установленные по окружности головы, регистрируют темп изменения радиоактивности, который пропорционален величине MK. Новейшей методикой исследования MK является позитронная эмиссионная томография (ПЭТ); в сочетании с применением короткоживущих изотопов 11 C и 15 O ПЭТ позволяет измерить потребление мозгом глюкозы и кислорода соответственно. ПЭТ подтвердила полученные другими методами данные, что регионарный мозговой кровоток (рМК) изменяется прямо пропорционально интенсивности метаболизма и варьируется от 10 до 300 мл/100 г/ мин. Например, при движениях в конечности быстро возрастает рМК в соответствующем участке двигательной коры. Аналогичным образом активизация зрения приводит к увеличению рМК в зрительной коре затылочных долей мозга.
Хотя MK равен в среднем 50 мл/100 г/мин, в сером веществе головного мозга он достигает 80мл/100 г/мин, в белом веществе — 20 мл/100 г/мин. MK у взрослых в среднем составляет 750 мл/мин, что соответствует 15-20 % от сердечного выброса. При MK ниже 20-25 мл/100 г/мин возникает повреждение мозга, что на ЭЭГ проявляется замедлением ритма. MK в пределах 15-20 мл/100 г/мин сответствует изоэлектрической линии на ЭЭГ, а при уменьшении MK до 10 мл/100 г/мин наступает необратимое повреждение мозга.
РЕГУЛЯЦИЯ МОЗГОВОГО КРОВООБРАЩЕНИЯ
1. Церебральное перфузионное давление
Церебральное перфузионное давление (ЦПД) —
это разница между средним артериальным давлением (АДср) и ВЧД (или церебральным венозным давлением). Если церебральное венозное давление значительно превышает ВЧД, то ЦПД равно разнице между АДср и церебральным венозным давлением. В физиологических условиях ВЧД незначительно отличается от церебрального венозного давления, поэтому принято считать, что ЦПД = = АДср - ВЧД. В норме церебральное перфузионное давление составляет 100 мм рт. ст. и зависит главным образом от АДср, потому что ВЧД у здорового человека не превышает 10 мм рт. ст.
При выраженной внутричерепной гипертензии (ВЧД > 30 мм рт. ст.) ЦПД и MK могут значительно снижаться даже при нормальном АДср. ЦПД < 50 мм рт. ст. проявляется замедлением ритма на ЭЭГ, ЦПД в пределах от 25 до 40 мм рт. ст. — изолинией на ЭЭГ, а при устойчивом снижении ЦПД менее 25 мм рт. ст. возникает необратимое повреждение мозга.
2. Ауторегуляция мозгового кровообращения
В головном мозге, так же как в сердце и почках, даже значительные колебания АД не оказывают существенного влияния на кровоток. Сосуды мозга быстро реагируют на изменение ЦПД. Снижение ЦПД вызывает вазодилатацию сосудов мозга, увеличение ЦПД — вазоконстрикцию. У здоровых людей MK остается неизменным при колебаниях АДср в пределах от 60 до 160 мм рт. ст. (рис. 25-1). Если АДср выходит за границы этих значений, то ауторегуляция MK нарушается. Увеличение АДср до 160 мм рт. ст. и выше вызывает повреждение ге-матоэнцефалического барьера (см. ниже), чреватое отеком мозга и геморрагическим инсультом. хронической артериальной гипертонии кривая ауторегуляции мозгового кровообращения (рис. 25-1) смещается вправо, причем сдвиг затрагивает и нижнюю, и верхнюю границы. При артериальной гипертонии снижение АД до обычных значений (меньше измененной нижней границы) приводит к уменьшению MK, в то время как высокое АД не вызывает повреждения мозга. Длительная гипотензивная терапия может восстановить ауторегуляцию мозгового кровообращения в физиологических границах.
Существуют две теории ауторегуляции мозгового кровообращения — миогенная и метаболическая. Миогенная теория объясняет механизм ауторегуляции способностью гладкомышечных клеток церебральных артериол сокращаться и расслабляться в зависимости от АДср. Согласно метаболической теории, тонус церебральных артериол зависит от потребности мозга в энергетических субстратах. Когда потребность мозга в энергетических субстратах превышает их доставку, в кровь выделяются тканевые метаболиты, которые вызывают церебральную вазодилатацию и увеличение MK. Этот механизм опосредуют ионы водорода (их роль в церебральной вазодилатации описана раньше), а также другие вещества — оксид азот (NO), аденозин, простагландины и, возможно, градиенты ионной концентрации.
3. Внешние факторы
Парциальное давление CO2 и O2 в крови
Парциальное давление CO2 в артериальной крови (PaCO2 ) — наиболее важный внешний фактор, влияющий на MK. MK прямо пропорционален PaCO2 в пределах от 20 до ЗОммрт. ст. (рис. 25-2). Увеличение PaCO2 на 1 мм рт. ст. влечет за собой мгновен ное повышение MK на 1-2 мл/100 г/мин, уменьшение PaCO2 приводит к эквивалентному снижению MK. Этот эффект опосредуется через рН цереброспинальной жидкости и вещества мозга. Поскольку CO2 , в отличие от ионов, легко проникает, через гематоэнцефалический барьер, то на MK влияет именно острое изменение PaCO2 , а не концентрации HCO3 '. Через 24-48 ч после начала гипо- или гиперкапнии развивается компенсаторное изменение концентрации HCO3 " в спинномозговой жидкости. При выраженной гипервентиляции (PaCO2 < 20 мм рт. ст.) даже у здоровых людей на ЭЭГ появляется картина, аналогичная таковой при повреждении головного мозга. Острый метаболический ацидоз не оказывает значительного влияния на MK, потому что ион водорода (H+ ) плохо проникает через гематоэнцефалический барьер. Что касается PaO2 , то на MK оказывают воздействие только его значительные изменения. В то время как гипероксия снижает MK не более чем на 10 %, при тяжелой гипоксии (PaO2 < 50 мм рт. ст.) MK увеличивается в гораздо большей степени (рис. 25-2).
Температура тела
Изменение MK составляет 5-7 % на 1 0 C. Гипотермия снижает CMRO2 и MK, в то время как гипер-термия оказывает обратный эффект. Уже при 20 0 C на ЭЭГ регистрируют изолинию, но дальнейшее уменьшение температуры позволяет еще сильнее снизить потребление кислорода мозгом. При температуре выше 42 0 C потребление кислорода мозгом также снижается, что, по-видимому, обусловлено повреждением нейронов.
Вязкость крови
У здоровых людей вязкость крови не оказываетзначительного влияния на MK.
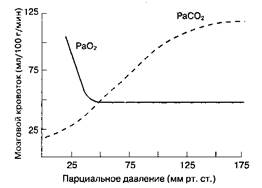
Рис. 25-2. Влияние PaO2 и PaCO2 Ha мозговой кровоток
Вязкость кровив наибольшей степени зависит от гематокрита, поэтому снижение гематокрита уменьшает вязкость и увеличивает MK. К сожалению, помимо этого благоприятного эффекта, снижение гематокрита имеет и отрицательную сторону: оно уменьшает кислородную емкость крови и, соответственно, доставку кислорода. Высокий гематокрит, например при тяжелой полицитемии, увеличивает вязкость крови и снижает MK. Исследования показали, что для лучшей доставки кислорода к мозгу гематокрит должен составлять 30-34 %.
Вегетативная нервная система
Внутричерепные сосуды иннервируются симпатическими (вазоконстрикторными), парасимпатическими (вазодилатирующими) и нехолинергическими неадренергическими волокнами; нейротрансмитте-ры в последней группе волокон — серотонин и вазо-активный интестинальный пептид. Функция вегетативных волокон сосудов мозга в физиологических условиях неизвестна, но продемонстрировано их участие при некоторых патологических состояниях. Так, импульсация по симпатическим волокнам pis верхних симпатических ганглиев может значительно сузить крупные мозговые сосуды и уменьшить MK. Вегетативная иннервация мозговых сосудов играет важную роль в возникновении церебрального вазоспазма после Ч MT и инсульта.
Гематоэнцефалический барьер
Между эндотелиальными клетками мозговых сосудов практически отсутствуют поры. Малочисленность пор — основная морфологическая особенность гематоэнцефалического барьера. Липидный барьер проницаем для жирорастворимых веществ, но значительно ограничивает проникновение ионизированных частиц и крупных молекул. Таким образом, проницаемость гематоэнцефалического барьера для молекулы какого-либо вещества зависит от ее размера, заряда, липо-фильности и степени связывания с белками крови. Углекислый газ, кислород и липофильные вещества (к которым относят большинство анестетиков) легко проходят через гематоэнцефалический барьер, в то время как для большинства ионов, белков и крупных молекул (например, маннитола) он практически непроницаем.
Вода свободно проникает через гематоэнцефалический барьер по механизму объемного тока, а перемещение даже небольших ионов затруднено (время полу выравнивания для натрия составляет 2-4 ч). В результате быстрые изменения концентрации электролитов плазмы (а значит, и осмолярности) вызывают преходящий осмотический градиент между плазмой и мозгом. Остро возникшая гипер-тоничность плазмы приводит к перемещению воды из вещества мозга в кровь. При острой гипотонич-ности плазмы, наоборот, происходит перемещение воды из крови в вещество мозга. Чаще всего равновесие восстанавливается без особых последствий, но в ряде случаев существует опасность быстро развивающихся массивных перемещений жидкости, чреватых повреждением мозга. Следовательно, значительные нарушения концентрации натрия или глюкозы в плазме нужно устранять медленно (см. гл. 28). Маннитол, осмотически активное вещество, которое в физиологических условиях не пересекает гематоэнцефалический барьер, вызывает устойчивое уменьшение содержания воды в мозге и часто используется для уменьшения объема мозга.
Целостность гематоэнцефалического барьера нарушают тяжелая артериальная гипертензия, опухоли мозга, ЧМТ, инсульт, инфекции, выраженная гиперкапния, гипоксия, устойчивая судорожная активность. При этих состояниях перемещение жидкости через гематоэнцефалический барьер определяется не осмотическим градиентом, а гидростатическими силами.
Цереброспинальная жидкость
Цереброспинальная жидкость находится в желудочках и цистернах головного мозга, а также в суб-арахноидальном пространстве ЦНС. Главная функция цереброспинальной жидкости — защита мозга от травмы.
Большая часть цереброспинальной жидкости вырабатывается в сосудистых сплетениях желудочков мозга (преимущественно в боковых). Некоторое количество образуется непосредственно в клетках эпендимы желудочков, а совсем небольшая часть — из жидкости, просачивающейся через периваскулярное пространство сосудов мозга (утечка через гематоэнцефалический барьер). У взрослых образуется 500 мл цереброспинальной жидкости в сутки (21 мл/ч), в то время как объем цереброспинальной жидкости составляет только 150 мл. Из боковых желудочков цереброспинальная жидкость через межжелудочковые отверстия (отверстия Монро) проникает в третий желудочек, откуда через водопровод мозга (сильвиев водопровод) попадает в четвертый желудочек. Из четвертого желудочка через срединную апертуру (отверстие Мажанди) и боковые апертуры (отверстия Люшка) цереброспинальная жидкость поступает в мозжечково-мозговую (большую) цистерну (рис. 25-3), а оттуда — в субарахноидальное пространство головного и спинного мозга, где и цир кулирует до тех пор, пока не всасывается в грануляциях паутинной оболочки больших полушарий. Для образования цереброспинальной жидкости необходима активная секреция натрия в сосудистых сплетениях. Цереброспинальная жидкость изото-нична плазме, несмотря на более низкую концентрацию калия, бикарбоната и глюкозы. Белок поступает в цереброспинальную жидкость только из перивас-кулярных пространств, поэтому его концентрация очень невелика. Ингибиторы карбоангидразы (аце-тазоламид), кортикостероиды, спиронолактон, фу-росемид, изофлюран и вазоконстрикторы уменьшают выработку цереброспинальной жидкости.
Цереброспинальная жидкость всасывается в грануляциях паутинной оболочки, откуда попадает в венозные синусы. Небольшое количество всасывается через лимфатические сосуды мозговых оболочек и периневральные муфты. Обнаружено, что всасывание прямо пропорционально ВЧД и обратно пропорционально церебральному венозному давлению; механизм этого явления неясен. Поскольку в головном и спинном мозге нет лимфатических сосудов, всасывание цереброспинальной жидкости — основной путь возвращения белка из интерстициальных и периваскулярных пространств мозга обратно в кровь.
Внутричерепное давление
Череп представляет собой жесткий футляр с нерастягивающимися стенками. Объем полости черепа неизменен, его занимает вещество мозга (80 %), кровь (12 %) и цереброспинальная жидкость (8 %). Увеличение объема одного компонента влечет за собой равное по величине уменьшение остальных, так что ВЧД не повышается. ВЧД измеряют с помощью датчиков, установленных в боковом желудочке или на поверхности полушарий головного мозга; в норме его величина не превышает 10 мм рт. ст. Давление цереброспинальной жидкости, измеренное при люмбальной пункции в положении больного лежа на боку, достаточно точно соответствует величине ВЧД, полученной с помощью внутричерепных датчиков.
Растяжимость внутричерепной системы определяют, измеряя прирост ВЧД при увеличении внутричерепного объема. Вначале увеличение внутричерепного объема хорошо компенсируется (рис. 25-4), но после достижения определенной точки ВЧД резко возрастает. Основные компенсаторные механизмы включают: (1) смещение цереброспинальной жидкости из полости черепа в субарахноидальное пространство спинного мозга; (2) увеличение всасывания цереброспинальной жидкости; (3) уменьшение образования цереброспинальной жидкости; (4) уменьшение внутричерепного объема крови (главным образом за счет венозной).
Податливость внутричерепной системы неодинакова в разных участках мозга, на нее влияют АД и PaCO2 . При повышении АД механизмы ауторе-гуляции вызывают вазоконстрикцию сосудов мозга и снижение внутричерепного объема крови. Артериальная гипотония, наоборот, приводит к ва-зодилатации сосудов мозга и увеличению внутричерепного объема крови. Таким образом, благодаря ауторегуляции просвета сосудов MK не изменяется при колебаниях АД. При повышении PaCO2 на 1 мм рт. ст. внутричерепной объем крови увеличивается на 0,04 мл/100 г.
Концепцию растяжимости внутричерепной системы широко используют в клинической практике. Растяжимость измеряют при введении стерильного физиологического раствора во внутрижелудоч-ковый катетер. Если после инъекций 1 мл раствора ВЧД увеличивается более чем на 4 мм рт. ст., то растяжимость считают значительно сниженной. Снижение растяжимости свидельствует об истощении механизмов компенсации и служит прогностическим фактором уменьшения MK при дальнейшем прогрессировании внутричерепной гипертензии. Устойчивое повышение ВЧД может вызвать катастрофическую дислокацию и вклинение различных участков мозга. Выделяют следующие виды повреждений (рис. 25-5): (1) ущемление поясной извилины серпом мозга; (2) ущемление крючка наметом мозжечка; (3) сдавленна продолговатого мозга при вклинении миндалин мозжечка в большое затылочное отверстие; (4) выпячивание вещества мозга через дефект черепа.
ВЛИЯНИЕ АНЕСТЕТИКОВ И ВСПОМОГАТЕЛЬНЫХ СРЕДСТВ НА ЦНС
Подавляющее большинство общих анестетиков благоприятно воздействует на ЦНС, снижая биоэлектрическую активность мозга. Катаболизм углеводов уменьшается, в то время как запасы энергии в виде АТФ, АДФ и фосфокреатина возрастают. Оценить эффект отдельного препарата очень сложно, потому что на него накладывается действие других средств, хирургическая стимуляция, растяжимость внутричерепной системы, АД и PaCO2 . Например, гипокапния и предварительное введение тиопентала предотвращают увеличение MK и ВЧД при использовании кетамина pi ингаляционных анестетиков. В этом разделе описано действие каждого препарата в отдельности. Итоговая табл. 25-1 позволяет оценить и сравнить влияние анестетиков и вспомогательных средств на ЦНС. В разделе также обсуждается роль миорелаксантов и средств, оказывающих воздействие на сосудистый тонус.
Ингаляционные анестетики
1. Испаряемые анестетики Метаболизм мозга
Галотан, энфлюран, десфлюран, севофлюран и изофлюран вызывают дозозависимое уменьшение метаболических потребностей мозга. Наибольшее снижение происходит при использовании изофлю-рана и энфлюрана (до 50 %), в то время как эффект галотана выражен слабее (потребность мозга в кислороде уменьшается не более чем на 25 %). Влияние десфлюрана и севофлюрана сходно с таковым у изофлюрана и энфлюрана. После того как на ЭЭГ регистрируется изолиния, дальнейшее увеличение дозы ингаляционного анестетика (в отличие от воздействия низких температур) не приводит к уменьшершю метаболических потребностей мозга. Более того, эффект анестетиков неодинаков в различных участках мозга: изофлюран угнетает метаболизм в основном в неокортексе. Энфлюран может провоцировать эпилептиформную активность на ЭЭГ, в этом случае метаболические потребности мозга значительно возрастают.
MK и внутричерепной объем крови
Ингаляционные анестетики вызывают дозозави-симую вазодилатацию мозговых сосудов и нарушение ауторегуляции мозгового кровообращения (рис. 25-6). Наиболее выраженное влияние на MK свойственно галотану: в дозе > 1 % он почти пол- ностью нарушает ауторегуляцию мозгового кровообращения. При использовании галотана MK увеличивается во всех отделах мозга. При равных МАК и АД галотан повышает MK на 200 %, а энфлюран и изофлюран — на 40 % и 20 % соответственно. В отличие от галотана, изофлюран увеличивает MK главным образом в субкортикальных участках больших полушарий, мозжечке, мосте и продолговатом мозге. По влиянию на MK десфлюран и севофлюран качественно и количественно напоминают изофлюран. Влияние ингаляционных анестетиков на MK зависит от длительности их применения: после 2-5-часовой ингаляции вышеперечисленных препаратов MK постепенно возвращается к норме.
Ингаляционные анестетики, как правило, не изменяют
8-09-2015, 23:29