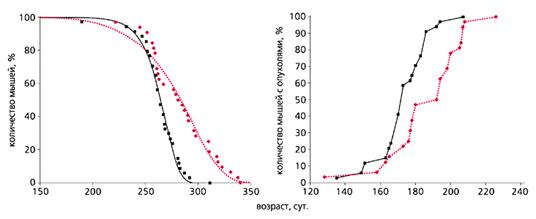
Рис. 3. Влияние метформина (цветная кривая) на продолжительность жизни
и развитие спонтанных опухолей (справа) у самок трансгенных мышей HER-2/neu [18].
Стареющие клетки: хорошие граждане, но плохие соседи
Не обольщайся призраком покоя: Бывает жизнь обманчива на вид. Настанет час, и утро роковое Твои мечты, сверкая, ослепит. Николай Заболоцкий |
В 1971 г. научный cотрудник Института биохимической физики РАН А.М.Оловников, используя данные о принципах синтеза ДНК в клетках, предложил гипотезу маргинотомии, объясняющую механизм работы счетчика клеточных делений. Согласно его гипотезе, ДНК-полимераза при матричном синтезе полинуклеотидов полностью не воспроизводит линейную матрицу, и реплика всегда получается короче в ее начальной части. Таким образом, при каждом делении клетки ее ДНК укорачивается, что ограничивает пролиферативный потенциал клеток и, очевидно, служит “счетчиком” числа делений и, соответственно, продолжительности жизни клетки в культуре. Открытие в 1985 г. фермента теломеразы, достраивающего укороченную теломеру в половых клетках и клетках опухолей, обеспечивая их бессмертие, стало блестящим подтверждением гипотезы Оловникова.
В последние годы достигнут значительный прогресс в изучении роли теломер в старении. Дисфункция теломер (связана ли с укорочением, прямым повреждением или ассоциированным с теломерой дефектным белком) может приводить к трем последствиям: старению клетки, ее гибели (факторы, подавляющие неопластический процесс) или нестабильности генома, которое может способствовать злокачественной трансформации клетки.
Старение клетки рассматривают как один из ее защитных механизмов при дисфункции теломер, поскольку оно останавливает пролиферацию, тем самым блокируя канцерогенез (“хорошие граждане”) [16]. В течение жизни в организме накапливаются соматические мутации, некоторые из них могут инактивировать гены, участвующие в клеточном старении. Кроме того, потеря гетерозиготности и мутации в генах-супрессорах (р53 и Rb) и онкогенах (например, ras) встречаются даже в нормальных клетках. Другой защитный механизм клетки с дисфункцией теломер, в который вовлечен интактный р53, - апоптоз. В клетках, где накапливаются мутации в гене р53 или в компонентах его регуляции, развивается нестабильность генома, повышающая вероятность злокачественной трансформации. Потеря дистальных областей теломер связана с уменьшением пролиферативной жизни клеток как in vitro, так и in vivo. Анализ данных по укорочению теломер в 15 различных тканях человека показал, что в год теломеры теряют от 20 до 60 пар оснований.
В настоящее время активно развивается гипотеза о важной роли укорочения теломер и соответственно реактивации теломеразы при старении и канцерогенезе. В опухолях человека появление бессмертных клеток (иммортализация) почти всегда обусловлено подавлением гена каталитической субъединицы теломеразы (hTERT). Множественность механизмов, подавляющих или регулирующих активность этого фермента, может объяснить исключительную редкость спонтанной иммортализации нормальных клеток человека. С другой стороны, введение субъединицы hTERT стимулирует рост различных типов клеток человека без последующего развития опухолевого роста. Вероятно, основная функция супрессорного гена p53 состоит в остановке роста в ответ на потерю теломер в старых клетках. Этой гипотезе не противоречат данные о поведении большинства опухолей, в которых этот ген мутирован, и объясняют характеристики редких типов опухолей, в которых функция p53 сохранена.
Недавно показано, что in vitro старые фибробласты человека стимулируют пролиферацию предраковых и злокачественных эпителиальных клеток, которые способны образовывать опухоли при прививке голым мышам (“плохие соседи”) [17]. У фибробластов на более ранних пассажах (пресенильных) эта способность менее выражена. Несмотря на довольно убедительные аргументы в пользу связи между старением клеток, биологией теломеры и раком человека, данные о роли клеточного старения в старении человека довольно противоречивы и требуют дальнейших исследований.
Увеличение продолжительности жизни и риск рака
Все равно, по какой дороге идти. Куда-нибудь ты наверняка попадешь, если будешь идти достаточно долго. Льюис Кэррол. Приключения Алисы в Стране чудес |
Расчеты показывают, что у человека с продолжительностью жизни 70 лет, крысы (2.5-3 года) и мыши (2-2.5 года) частота новообразований одинакова и составляет 30% [3, 19]. Однако при нормализации по количеству клеток в организме оказывается, что мышь более склонна к развитию опухолей, чем человек. Этот феномен склонны объяснять тем, что в соматических клетках мыши теломераза более активна, чем в клетках человека, чему соответствует значительно большая длина теломер у мыши по сравнению с человеком. Вместе с тем видовая продолжительность жизни млекопитающих соотносится с эффективностью репарации ДНК и устойчивостью их клеток к окислительному стрессу. Так, в ДНК человека восстановление гуанина, алкилированного канцерогенными нитрососоединениями, в сотни раз выше, чем у мыши, что связано с большей резистентностью человека к этим агентам. Оказалось, что эффективность репарации вызываемых канцерогеном повреждений в ДНК различных органов связана с продолжительностью жизни мышей. Анализ данных о частоте рака у генетически модифицированных животных с увеличенной продолжительностью жизни говорит о снижении у них частоты злокачественных новообразований [3].
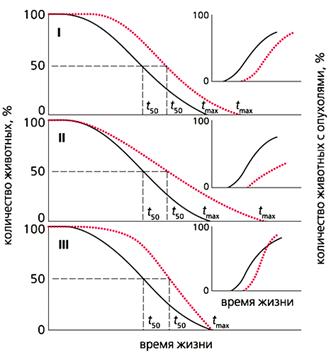
Рис. 4. Влияние трех типов геропротекторов (цветные кривые) на продолжительность жизни животных
и частоту спонтанных опухолей (на врезке).
I - продолжительность жизни всех особей увеличивается, скорость старения не меняется, новообразования развиваются позднее, чем в контрольной группе, но с такой же частотой;
II - скорость старения снижается, частота новообразований уменьшается;
III - продолжительность жизни не меняется, а скорость старения и количество новообразований растет.
tmax - максимальная продолжительность жизни, t50 - время полужизни.
Показано, что различные геропротекторы по-разному влияют на развитие новообразований, что определяется в основном типом замедления старения в популяции (рис.4). Наши наблюдения свидетельствуют, что более “прямоугольный” характер кривых выживания ассоциирован с увеличенной скоростью развития фатальных опухолей у крыс.
С конца ХVIII в. и вплоть до середины ХХ в. в экономически развитых странах постепенно увеличивалась частота раковых заболеваний. Частота всех злокачественных новообразований в США и многих других странах, включая Россию, заметно росла с 1960 по 1990 г. и у мужчин, и у женщин, что совпадало с увеличением продолжительности жизни. С 90-х годов ХХ в. в наиболее экономически развитых странах, таких как США, Швеция и Дания, частота рака стала несколько снижаться (рис.5). И именно во второй половине ХХ в. в наиболее развитых странах изменился характер траекторий смертности, что сопровождалось уменьшением смертности в самых старших возрастах (рис.6). В то же время в других странах, например в России, частота злокачественных новообразований продолжает расти. Очевидно, что увеличение продолжительности жизни и характера траектории смертности по типу параллельного сдвига или даже увеличения ее наклона будет способствовать снижению частоты злокачественных новообразований.
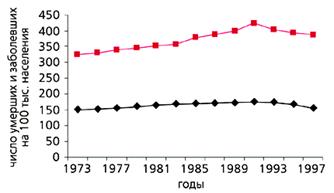
Рис. 5. Динамика заболеваемости (цветная кривая) и смертности от рака в США в XX в. [19].
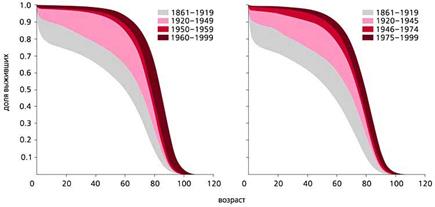
Рис. 6. Динамика изменений выживаемости женщин (слева) и мужчин в Швеции с 1861 по 1999 г. [20].
* * *
Наш великий соотечественник И.И.Мечников - основоположник современной геронтологии и автор самого термина “геронтология” - весной 1907 г. в своем предисловии к первому изданию “Этюдов оптимизма” писал: “Наука в России переживает продолжительный и тяжелый кризис. На науку не только нет спроса, но она находится в полнейшем загоне”. К сожалению, сегодня в России ситуация мало чем отличается от той, что была 100 лет назад. Бюджет всей российской науки не превышает бюджета Национального института старения США - одного из самых “маленьких” в системе Национального института здоровья, и существенно меньше, чем средства, выделяемые государством на содержание чиновников. Однако стареют и они.
И к ним, а также нашим депутатам, мы адресуем Открытое письмо, опубликованное недавно в Интернете (http://cureaging.com; http://ieet.org/index.php/IEET/ more /scientistsantiaging/; http://www.longevity-science.blogspot.com/ и др.), подписанное 54 ведущими учеными-геронтологами мира. Ниже приводится перевод этого письма на русский язык:
“Всем, кого это касается.
На моделях многих сильно отличающихся друг от друга видов лабораторных животных (нематоды, дрозофилы, мыши и т.п.) удается достигнуть замедления старения и продления активной жизни. Поэтому, исходя из общности фундаментальных механизмов старения, есть основания полагать возможным замедление старения у людей.
Расширение наших знаний о старении позволит лучше противостоять таким истощающим организм патологиям, связанным со старением, как рак, сердечно-сосудистые заболевания, диабет II типа и болезнь Альцгеймера. Терапия, основанная на знании фундаментальных механизмов старения, будет способствовать лучшему противодействию этим возрастным патологиям.
Именно поэтому наше письмо является призывом к увеличению финансирования этой области, что крайне необходимо для интенсификации как исследований фундаментальных механизмов старения, так и поиска способов его замедления. Все это может привести к намного большим дивидендам, чем в случае, если бы эти же средства и усилия вкладывались в непосредственное противостояние возрастным патологиям. Поскольку механизмы старения становятся все более и более понятными, могут быть разработаны эффективные средства вмешательства в этот процесс. Это позволит значительному количеству людей продлить здоровую и продуктивную жизнь”.
Список литературы
1. Oeppen J., Vaupel J.W. // Science. 2002. V.296. P.1029-1031.
2. Старение населения Санкт-Петербурга: социально-демографические аспекты / Ред. Г.Л.Сафарова. СПб., 2006.
3. Анисимов В.Н. Молекулярные и физиологические механизмы старения. СПб., 2003.
4. Handbook of the Biology of Aging. Sixth Edition / Eds E.J.Masoro, S.N.Austad. Amsterdam, 2006.
5. Cutler Mattson // Aging Cell. 2006. V.6.
6. Perls T., Levenson R., Regan M., Puca A. // Mech. Ageing Dev. 2002. V.123. P.231-242.
7. Westendorp R.G., Kirkwood T.B.L. // Nature.1998. V.396. P.743-746.
8. Orr W.C., Sohal R.S. // Science. 1994. V.263. P.1128-1130.
9. Parkes T.L., Elia A.J., Dickinson D. et al. // Nature Genetics. 1998. V.19. P.171-174.
10. Tatar M., Bartke A., Antebi A. // Science. 2003. V.299. P.1346-1351.
11. Lin Y.J., Seroude L., Benzer S. // Science.1998. V.282. P.943-946.
12. Coschigano K.T., Clemmons D., Bellush L.L., Kopchick J.J. // Endocrinology. 2000. V.141. P.2608-2613.
13. Bartke A., Coschigano K., Kopchick J. et al. // J. Gerontol. Biol. Sci. 2001. V.56A. P.B340-B349.
14. Holzenberger M., Dupond J., Ducos B. et al. // Nature. 2003. V.421. P.182-187.
15. Fontana L., Meyer T.E., Klein S., Holloszy J.O. // Proc. Natl. Acad. Sci. USA. 2004. V.101. P.6659-6663.
16. Anisimov V.N., Berstein L.M., Egormin P.A. et al. // Exp. Gerontol. 2005. V.40. P.685-693.
17. Krtolica A., Campisi J. // Успехи геронтол. 2003. Т.11. С.109-116.
18. Anisimov V.N., Ukraintseva S.V., Yashin A.I. // Nature Rev. Cancer. 2005. V.5. P.807-819.
19. Ries L.A.G., Eisner M.P., Kosary C.L. et al. // SEER Cancer Statistics Review 1973-1998. Nat. Can. Institute. Bethesola. MD. 2001.
20. Vashin A.I., Begun S.S., Boikos I. et al. // Mech. Ageing Dev. 2002. V.123. P.637-647.
8-09-2015, 21:41