СОДЕРЖАНИЕ
Введение
1. Активное наступление сна или лишение бодрствования?
2. Стадии медленного сна и быстрый сон
3. Сон в онто- и филогенезе
4. Потребность в сне
5. Депривация сна
6. Сновидения
Вывод
Список литературы
ВВЕДЕНИЕ
Сон - это периодическое физиологическое состояние мозга и организма человека и высших животных, внешне характеризующееся значительной обездвиженностью и отключением от раздражителей внешнего мира.
Субъективно у человека при этом угнетается сознаваемая психическая активность, периодически восстанавливающаяся при переживании сновидений, часто с последующим их забыванием. Низшим животным свойственны периоды бездеятельного состояния. Однако еще не выяснено, в какой мере оно функционально соответствует сну высших животных. У человека на сон уходит примерно треть отпущенного ему времени на жизнь. Когда он спит, ему присущи сновидения - субъективно переживаемые психические явления, периодически возникающие во время естественного сна. Интерес к сновидениям характерен для всех эпох человеческой культуры, однако подход к ним существенно менялся на протяжении истории.
Человек видит во сне то, что когда-то увидено, задумано, воспринято мозгом, оставило свой, пусть мимолетный, след в нервных клетках мозга. Хорошо известно, что слепым от рождения не снятся зрительные образы. Иначе говоря, во сне можно увидеть только то, что было. Но в каком виде? Человек видит порой совершенно сказочные, невероятные сновидения. Чего только не бывает во сне! Мы видим себя в далеком детстве, путешествуем по различным странам, сражаемся, без удивления встречаемся с умершими людьми, говорим с животными, как в сказках, летаем по воздуху. В мозгу спящего, как в кино, за короткое время проходит порой вся человеческая жизнь. И какие бы фантастические картины ни развертывались во сне, все они кажутся подлинными, реальными.
1 АКТИВНОЕ НАСТУПЛЕНИЕ СНА ИЛИ ЛИШЕНИЕ БОДРСТВОВАНИЯ?
Уже в самых ранних исследованиях механизмов сна четко намечаются две основные точки зрения на эту проблему. Первая — сон возникает в результате активного процесса, возбуждения определенных структур («центров сна»), которое вызывает общее снижение функций организма (активные теории сна). Вторая - это пассивные теории сна, или теории деафферентации, согласно которым сон наступает пассивно в результате прекращения действия каких-то факторов, необходимых для поддержания бодрствования. Различия между этими направлениями удачно определил Н. Клейтман, писавший, что «заснуть» и «не суметь остаться бодрствующим», - это не одно и то же, так как первое предполагает активное действие, а второе — пассивное устранение активного состояния.
В первых неврогенных теориях сна, возникших в конце прошлого и начале нынешнего столетия в результате клинических наблюдений после эпидемии летаргического энцефалита, отражаются эти две противоположные тенденции. Так, Л. Маутнер пришел к выводу, что причиной наступления сна при этом заболевании была опухоль в области среднего мозга, вызывавшая сдавление афферентных путей и прекращение потока сенсорных импульсов в высшие отделы мозга. Фон Экономо же считал, что сон наступает в результате торможения таламуса и коры больших полушарий «центром сна». Исходя из противоположного характера ведущих симптомов заболевания (сонливости или бессонницы) он полагал, что центр сна состоит, по крайней мере, из двух частей — каудальной части, являющейся как бы центром бодрствования, и ростральной, действующей как центр сна.
Первыми экспериментальными исследованиями, свидетельствующими о существовании центра сна, явились работы В. Гесса. Показав, что слабое электрическое раздражение четко ограниченной области промежуточного мозга у подопытных кошек вызывало сон со всеми подготовительными фазами (потягивание кошки, умывание, принятие характерной позы), В. Гесс высказал предположение, что существует центр, возбуждение которого обеспечивает наступление естественного сна. В дальнейшем опыты В.Гесса были подтверждены многочисленными исследователями, которые вызывали наступление сна у подопытных животных при помощи электрического и химического раздражения гипоталамуса и прилежащих структур, и теория о центре сна получила значительное признание.
Однако против подобного локализационистского объяснения механизма возникновения сна решительно выступал И.П. Павлов. Он рассматривал сон как результат торможения коры больших полушарий; при этом его теория сна не исключала участия и подкорковых структур в возникновении сна. В качестве основных этапов в дальнейшем развитии «пассивных» теорий можно отметить работы Н. Клейтмана, который пришел к выводу о том, что сон — это «легко обратимая пассивность высших функциональных центров коры», которая наступает в результате уменьшения потока афферентной импульсации. Эксперименты Ф. Бремера с поперечными перерезками на разных уровнях ствола мозга кошки, показавшие, что межколликулярная перерезка приводит к картине сна на ЭЭГ (спящий «cerveauisole»), тогда как перерезка на границе головного и спинного мозга не изменяет картину бодрствования на ЭЭГ (бодрствующий «encephaleisole»), подтверждали представление о сне как результате деафферентации коры больших полушарий. В результате опытов с разрушением различных областей промежуточного мозга у обезьян, приводивших к наступлению сна, С. Рэнсон пришел к выводу о том, что гипоталамус является центром «интеграции эмоционального выражения» и сон наступает в результате периодического снижения активности этого центра бодрствования.
Открытие Дж. Моруцци и X. Мэгуном в 1949 г. восходящего активирующего влияния неспецифической ретикулярной системы (ВРАС) значительно усилило позиции пассивных теорий сна. Поддержание бодрствующего состояния объяснялось теперь тоническим влиянием ВРАС. Дальнейшие исследования привели к открытию и других активирующих систем — диффузной и специфической таламической систем и активирующих структур заднего гипоталамуса.
Впоследствии в целом ряде работ было показано, что раздражение ретикулярных структур ствола мозга может вызывать не только пробуждение, но и сон. Так, низкочастотное раздражение почти всех исследуемых пунктов ретикулярной формации (область среднего мозга, моста и продолговатого мозга) у кошек вызывало наступление сна, что позволило Дж. Росси и А. Цанкетти предположить существование в стволе мозга двух антагонистических механизмов — десинхронизирующего (пробуждающего) и синхронизирующего (вызывающего сон), — поэтому сон не пассивное состояние отсутствия бодрствования, а результат активного торможения деятельности мозга. Моруцци также привел экспериментальные доказательства того, что в нижних отделах ствола мозга имеются синхронизирующие структуры, действующие как антагонисты ВРАС. К синхронизирующим структурам, раздражение которых может вызвать наступление сна у животного, позже были отнесены и хвостатое ядро, и неспецифическая таламическая система, и структуры базальной части переднего мозга, вчастности, преоптическая область.
Одна из попыток создания единой теории сна была предпринята П.К. Анохиным. Состояние сна он представлял как результат проявления целостной деятельности организма, строго координирующей корковые и подкорковые структуры в единую функциональную систему. В своей гипотезе П.К. Анохин исходил из того, что гипоталамические «центры сна» находятся под тоническим угнетающим влиянием со стороны коры больших полушарий. Именно поэтому при ослаблении этого влияния вследствие снижения рабочего тонуса корковых клеток («активный сон» по Павлову) гипоталамические структуры как бы «высвобождаются» и определяют всю ту сложную картину перераспределения вегетативных компонентов, которая характерна для состояния сна. При этом гипоталамические центры оказывают угнетающее влияние на восходящую активирующую систему, прекращая доступ в кору всего комплекса активирующих воздействий (и наступает «пассивный сон» по Павлову). Эти взаимодействия представляются циклическими, поэтому состояние сна может быть вызвано искусственно (или в результате патологического процесса) воздействием на любую часть этого цикла (рис. 1.1).
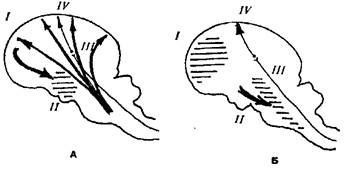
Рис. 1.1. Схема взаимодействия «центров сна» и «пробуждающих» структур при бодрствовании и наступлении сна (по П.К Анохину)
А. Бодрствование. Корковые влияния ( I ) тормозят «центры сна» ( ІІ ) и восходящие активирующие влияния ретикулярных структур ( ІІІ ) и возбуждения, идущие по лемнисковым путям ( IV ), свободно достигают коры
Б. Сон. Заторможенные отделы коры ( I ) перестают оказывать сдерживающие влияния на «центры сна» ( II ), и они блокируют восходящие активирующие влияния ( III ), не затрагивая возбуждений по лемнисковым путям ( IV ) .
В качестве иллюстрации этой гипотезы рассматривается сон новорожденного — поскольку в коре еще не имеется достаточно зрелых элементов для активного нисходящего тормозного влияния на уже значительно созревшие гипоталамические структуры, основным состоянием новорожденного является состояние сна. Причиной пробуждения является голод - возбуждаемые «голодной» кровью гипоталамические структуры реципрокно затормаживают «центры сна», устраняя их угнетающее влияние на активирующие системы, и ребенок просыпается. После насыщения восстанавливаются прежние соотношения, и ребенок снова засыпает.
2. СТАДИИ МЕДЛЕННОГО СНА И БЫСТРЫЙ СОН
Основные данные, полученные за годы многочисленных и разнообразных исследований сна, сводятся к следующему. Сон — не перерыв в деятельности мозга, это просто иное состояние. Во время сна мозг проходит через несколько различных фаз, или стадий, активности, повторяющейся с примерно полуторачасовой цикличностью. Сон состоит из двух качественно различных состояний, называемых медленным и быстрым сном. Они отличаются по суммарной электрической активности мозга (ЭЭГ), двигательной активности глаз (ЭОГ), тонусу мышц и многочисленным вегетативным показателям (частоте сердечных сокращений и дыхания, электрической активности кожи и т.д..
Медленный сон подразделяется на несколько стадий, выделенных на основании изменений ЭЭГ (рис. 2.1) и отличающихся по глубине. В первой стадии исчезает основной биоэлектрический ритм бодрствования — альфа-ритм. Он сменяется низкоамплитудными колебаниями различной частоты. Это стадия дремоты, засыпания. При этом у человека могут возникать сноподобные галлюцинации. Вторая стадия (поверхностный сон) характеризуется регулярным появлением веретенообразного ритма 14—18 колебаний в секунду («сонные» веретена). С появлением первых же веретен происходит отключение сознания; в паузы между веретенами человека легко разбудить. Третья и четвертая стадии объединяются под названием дельта-сна, потому что во время этих стадий на ЭЭГ появляются высокоамплитудные медленные волны — дельта-волны. В третьей стадии они занимают от 30 до 50% всей ЭЭГ. В четвертой стадии дельта-волны занимают более 50% всей ЭЭГ. Это наиболее глубокая стадия сна, здесь наивысший порог пробуждения, самое сильное отключение от внешнего мира. При пробуждении в этой стадии человек с трудом ориентируется, в наибольшей степени компрессирует время (недооценивает длительность предшествующего сна). Дельта-сон преобладает в первую половину ночи. При этом снижается мышечный тонус, становятся регулярными и урежаются дыхание и пульс, понижается температура тела (в среднем на 0,5о ), отсутствуют движения глаз, может регистрироваться спонтанная кожно-гальваническая реакция.
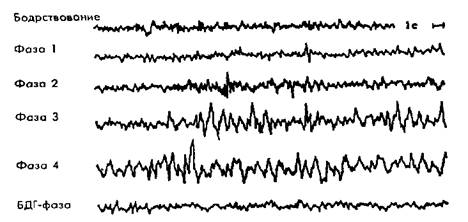
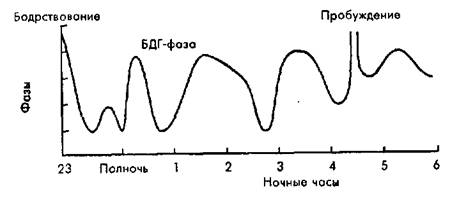
Рис. 2.1. Фазы сна. ЭЭГ при различных фазах сна (вверху). Изменения глубины сна на протяженииночи, удлинение периодов БДГ–сна(внизу).
Быстрый сон — самая последняя стадия в цикле сна. Она характеризуется быстрыми низкоамплитудными ритмами ЭЭГ, что делает ее похожей на ЭЭГ при бодрствовании. Усиливается мозговой кровоток, на фоне глубокого мышечного расслабления наблюдается мощная активация вегетатики. Помимо тонических компонентов стадии бістрого сна, выявляются фазические компоненты — быстрые движения глазных яблок при закрытых веках (БДГ, или КЕМ — rapideyemovements), мышечные подергивания в отдельных группах мышц, резкие изменения частоты сердечных сокращений (от тахикардии к брадикардии) и дыхания (серия частых вдохов-выдохов, потом пауза), эпизодические подъемы и падения кровяного давления, эрекция полового члена у мужчин и клитора у женщин. Порог пробуждения колеблется от высокого до низкого. Именно в этой стадии возникает большая часть запоминающихся сновидений. Синонимы быстрого сна — парадоксальный (активированный характер ЭЭГ при полной мышечной атонии), RЕМ,или БДГ–сон, ромбэнцефальный (в связи с локализацией регулирующих механизмов).
Весь ночной сон состоит из 4—5 циклов, каждый из которых начинается с первых стадий медленного и завершается быстрым сном. Каждый цикл продолжается около 90—100 мин. В двух первых циклахпреобладает дельта-сон, эпизоды быстрого сна относительно коротки. В последних циклах преобладает быстрый сон, а дельта-сон резко сокращен и может отсутствовать (рис. 2.1). В отличие от многих животных, человек не просыпается после каждого цикла сна. Структура сна у здоровых людей более или менее сходна — 1-я стадия занимает 5—10% сна, 2-я — 40—50%, дельта-сон — 20—25%, быстрый сон - 17-25%
Таким образом, каждую ночь 4—5 раз мы видим сны, и «разглядывание» сновидений занимает в общей сложности от 1 до 2 ч. Люди, утверждающие, что они видят сновидения очень редко, просто не просыпаются в фазе сновидений. Интенсивность самих сновидений, степень их необычности и эмоциональной насыщенности может быть различной, но факт их регулярного возникновения во время сна не вызывает сомнений.
Распространенное в прошлом представление о том, что сон необходим для «отдыха» нейронов головного мозга и характеризуется снижением их активности, исследованиями нейрональной активности не подтвердились. Во время сна в целом не происходит уменьшения средней частоты активности нейронов по сравнению с состоянием спокойного бодрствования. В быстром же сне спонтанная активность нейронов может быть выше, чем в напряженном бодрствовании. В медленном и быстром сне активность различных нейронов организована по-разному.
Кроме электрофизиологических, для отдельных стадий сна характерны определенные гормональные сдвиги. Так, во время дельта-сна увеличена секреция гормона роста, стимулирующего тканевой обмен. Во время быстрого сна усилена секреция гормонов коры надпочечников, которая в бодрствовании возрастает при стрессе. Интенсивность энергетического обмена в мозговой ткани во время медленного сна почти такая же, как в состоянии спокойного бодрствования, а во время быстрого сна значительно выше.
Таким образом, можно утверждать, что мозг активен во время сна, хотя эта активность качественно иная, чем при бодрствовании, и в разных стадиях сна имеет свою специфику.
3 СОН В ОНТО- И ФИЛОГЕНЕЗЕ
В онтогенезе соотношение «сон—бодрствование» изменяется. Так, у новорожденных состояние бодрствования составляет лишь незначительную часть суток, и значительную часть сна занимает быстрый сон. По мере взросления уменьшается общее количество сна, изменяется соотношение фаз внутри цикла сна — сокращается быстрый сон и относительно возрастает медленный сон, к 14 годам цикл сна достигает 90 мин. У взрослого человека, как уже говорилось, быстрый сон занимает около 1/4 всего времени сна. В старческом возрасте происходит уменьшение общего количества сна, при этом сокращается и медленный, и быстрый сон. После 75 лет часто наблюдается невротическая бессонница — редуцируется медленный сон, сон становится прерывистым, нарушаются циклы сна.
Чередование периодов активности и покоя происходит у всех живых существ; возможно, периоды покоя — это аналоги медленного сна. В том или ином виде сон наблюдается у всех позвоночных. Но сон, состоящий из нескольких циклоп, внутри которых развертываются стадии медленного и быстрого сна, присущ только теплокровным. По своей организации сон млекопитающих и птиц не отличается от сна человека, хотя медленный сон у животных менее дифференцирован, процентное соотношение медленного и быстрого сна различно у разных животных, а циклы сна, как правило, короче. «Короткая интенсивная жизнь идет рука об руку с долгим сном и коротким циклом сна». У крысы цикл сна продолжается 12 мин, у собаки — 30 мин, у слона — около 2 ч. Особенности организации сна связаны с экологией животных.
У птиц периоды быстрого сна очень короткие — при этом, в связи с полной мышечной атонией, опускается голова, падают крылья. Если птица сидит на ветке, то при падении тонуса мышц ног сжимаются пальцы, и птица может спать, не падая с ветки.
Сон копытных также связан с их образом жизни — стадностью, боязнью хищников — и имеет характер «рваного» сна (после каждого цикла сна животное поднимает голову и осматривается, поэтому в каждый данный момент некоторые особи обязательно бодрствуют). Характер растительной пищи требует длительного пережевывания, и поверхностные стадии сна протекают у жвачных во время жевания.
У норковых млекопитающих хорошо выражена цикличность, они много спят, и быстрый сон занимает до 1/3 всего времени сна. Для многих из них характерна сезонная спячка. Она характеризуется потерей способности к терморегуляции, резким уменьшением количества дыхательных движений и сердечных сокращений, падением общего уровня обмена веществ. У некоторых крупных млекопитающих (медведи, еноты, отчасти барсуки) наблюдается сезонный сон, или факультативная спячка. В этом случае температура тела, количество дыхательных движений и общий уровень обменных явлений снижаются мало. При изменении внешних условий подобный сон легко может быть прерван.
Особенности сна морских млекопитающих также связаны с их экологией. Для каждого дыхательного акта как во время сна, так и во время бодрствования они должны всплыть, чтобы выставить ноздри в воздух. В зависимости от образа жизни возникли разные формы адаптации. Так, при электрофизиологической регистрации сна у дельфинов Л. Мухаметов обнаружил феномен «однополушарного» сна — дельта-волны возникали только в одном полушарии (поочередно в правом или левом). При этом в другом полушарии картина ЭЭГ соответствовала поверхностным стадиям медленного сна или бодрствованию. ЭЭГ, соответствующая поверхностным стадиям медленного сна, могла наблюдаться и в обоих полушариях одновременно; признаков быстрого сна выявить не удалось. Такой же «однополушарный» медленный сон появляется у так называемых ушастых тюленей (котики и сивучи), когда они находятся в бассейне и не могут выйти на сушу. Когда же они спят на суше, в обоих полушариях у них наблюдается ЭЭГ, свойственная обычному медленному сну; регистрируется много эпизодов быстрого сна.
У тюленей и морских львов, которые только часть жизни проводят в воде, весь цикл сна развивается в течение дыхательной паузы. Они хорошо «продыхиваются», делая несколько глубоких вдохов, и ныряют. За 15—20 мин происходит смена стадий медленного сна и быстрый сон, и они выныривают для следующего «продыхивания».
Таким образом, сон является жизненно необходимым для высокоорганизованных животных. При этом особенности сна различных животных отражают его приспособительный характер к условиям обитания и факторам внешней среды.
4. ПОТРЕБНОСТЬ В СНЕ
Многие люди хотели бы спать меньше, так как сон, по их мнению, — это потерянное для жизни время. Другие, наоборот, хотели
9-09-2015, 18:37