После возникновения теории информации возрастание времени реакции при увеличении числа альтернатив стало связываться с возрастанием информации, подсчитьтаемой по известной формуле Шеннона. Такое возрастание времени реакции при возрастании числа альтернатив было названо законом Хика.
Однако дальнейшие исследования показали ограниченность дейст-2*
вия закона Хика. Было установлено, что уже после десяти сигналов время реакции перестает возрастать (314). 'Это значит, что уже в ус . ловиях реакции выбора при определенном количестве сигналов про. цесс активного выбора реакций сменяется процессом, имеющим существенно иной механизм.
Переводчик в эксперименте со случайной подачей иностранных слов может быть уподобен испытуемому в таком опыте с измерением времени реакций, в котором он должен быть готовым отреагировать одной из имеющихся у него нескольких сотен или тысяч реакций при возникновении одного из возможных нескольких сотен (тысяч) сигналов. Будет ли в данном случае время его речевой реакции перевода существенно, во много раз отличаться от времени двигательной реакции в условиях эксперимента с несколькими кнопками или сиг. налами?
Переводчик оперирует не только со словами, но и с речевыми структурами, грамматическими формами. Здесь возникает вопрос о временных параметрах актуализации грамматических форм в эксперименте, в котором требуется как можно быстрее опознать речевую структуру. Представляло интерес выяснить, какие существуют временные различия между актуализацией слов и актуализацией речевых структур.
Во многих исследованиях по педагогической психологии содержится идея свертывания и автоматизации операций при достижении человеком высшего уровня мастерства в том или ином виде деятельности. Согласно этой точке зрения, состав, язык операций, действий остается прежним. Речевая же деятельность синхронного переводчика позволяет предположить, что высший уровень овладения деятельностью предполагает качественно иной, волновой язык процесса, который именно на этом высшем уровне обнаруживает себя в полной мере.
Временные параметры речевых реакций переводчиков определялись в условиях лабораторного эксперимента, в котором в качестве испытуемых участвовали студенты П-Ш курсов иностранного факультета университета (исследование Д. В. Балубовой и др.) (310). Количество английских слов, входящих в активный словарь этой категории испытуемых, превышало одну тысячу. Последовательности из тридцати английских слов, которые предъявлялись испытуемым для перевода, составлялись на основе случайной выборки. Инструкция первой серии опытов требовала, чтобы при предъявлении английского слова испытуемый как можно скорее отвечал адекватным русским словом. Слова в первой серии предъявлялись в двух вариантах — на слух и для зрительного восприятия. Оба варианта записывались на магнитофон. Время речевой реакции перевода определялось с помощью измерения длины магнитной ленты, прошедшей между предъявлением слова и началом ответа испытуемого. Для каждого испытуемого подсчитывалось среднее время его реакции перевода по тридцати словам. Совокупность всех значений времени перевода была объектом анализа в первой серии.
Во второй серии экспериментов определялось время распознавания
матических структур. В опытах этой серии испытуемым предлага-английские фразы, состоящие из шести или восьми слов. Инструк-
требовала определить содержащуюся в этих фразах грамматическую Структуру за минимально короткое время и назвать вид этой структуры. $сего было три структуры. Фразы предлагались как для зрительного,
и для слухового восприятия. Фразы были составлены таким образом, что заканчивались на основном элементе грамматической структуры; это лишало испытуемого дополнительного времени обдумывания при зрительном или слуховом восприятии фразы.
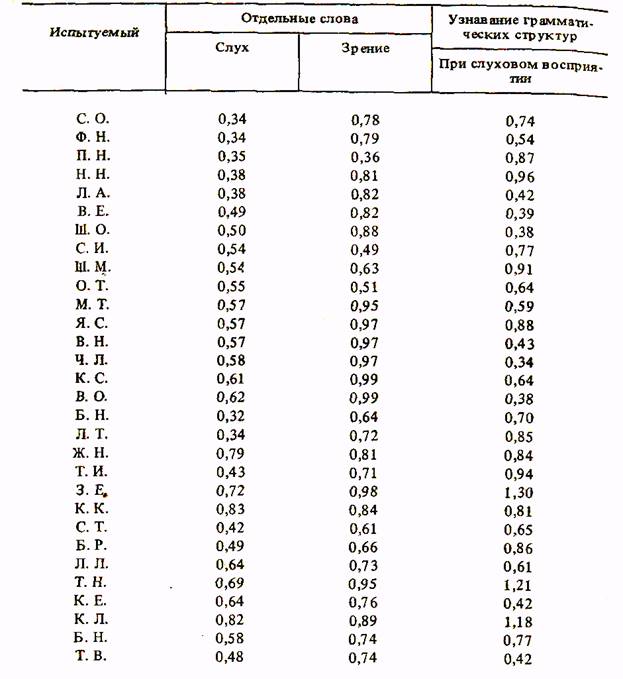
В таблице 1 приведены полученные в двух сериях эксперимента материалы. Как видно из таблицы, между показателями реакции перевода отдельных слов при зрительном и слуховом восприятии обнаруживается существенная разница: за исключением отдельных испытуемых, при слуховом восприятии слов у большинства время реакции короче, чем при восприятии зрительном. Это соответствует данным о времени зрительных и слуховых реакций (333).
Особенно резкие различия в скорости реакции между зрением и слухом были обнаружены во время распознавания грамматических структур. Реакция на английские фразы при зрительном их восприятии оказалась значительно длительнее, чем при слуховом восприятии. Поскольку эти различия связаны, по-видимому, с особенностями процесса чтения и физиологическими процессами, происходящими в зрительном анализаторе, а также в связи с тем, что нас интересует прежде всего центральное звено актуализации прошлого опыта, мы исключили время зрительных реакций на грамматические структуры и ограничились лишь рассмотрением реакций при слуховом восприятии.
Как уже говорилось, число распознаваемых грамматических структур в настоящем эксперименте равнялось трем. Поскольку первая структура могла длительнее анализироваться в силу ориентировки, а третья структура распознавалась слишком быстро, было решено при рассмотрении материала второй серии подвергнуть анализу показатели времени реакции лишь второй грамматической структуры при слуховом восприятии английских фраз.
Рассматривая таблицу 1, мы видим, что основное количество значений времени перевода отдельных слов приходится на показатели времени реакции меньше 0,4 с. Таких значений времени при слуховом восприятии слов оказалось 70%. 43% составили показатели, не превышающие 0,5 с, 23% — показатели меньше 0,4 с. Наименьший показатель был обнаружен у испытуемого Б. Н.—0,32 с. Что касается числа показателей, больших 0,7 с, то их при слуховом восприятии слов оказалось всего три случая, т. е. 10% показателей.
Как уже говорилось, в соответствии с литературными данными о времени реакций длительность реакций при зрительном восприятии слов в массе своей существенно превысила длительность реакций при
37
Таблица 1 Время актуализации английских слов и грамматических структур, с
слуховом восприятии. Однако ни у кого из наших испытуемых среднее время распознавания английских слов, предъявленных для зрительного восприятия, не достигло 1 с.
Количество показателей времени реакции при зрительном восприятии слов меньшее, чем 0,7 с, составило всего 23%. 50% показателей в этом варианте опыта составили значения, не достигшие 0,8 с, и 50% значений пришлось на величины времени в промежутке между 0,8 и 0,99 с.
38
Что же касается показателей времени реакции при слуховом рас->знавании грамматических структур, то значения этих показателей |*акже не превышали 1 с. Распределение времени реакции при распо-даавании грамматических структур похоже на распределение времени [йеакции при переводе отдельных слов на основе их зрительного вос-[яриятия. Также оказалось, что число значений, меньших 0,8 с, состави-фо 50%. 50% показателей составили те значения, которые были между ;0,8 и 0,99 с. Что же касается значений времени реакции, меньших 0,6 с, то таких значений оказалось всего три случая — 10%. Такова общая количественная характеристика времени реакций при переводе слов и грамматических структур, представленных в обобщающей таблице. Полученные экспериментальные данные приобретают смысл при сопоставлении их с известной в психологии длительностью простых психических процессов. Так, время прочтения слова, согласно исследованиям Кэтелла, близко к 0,43 с (333). Если сравнить полученное в наших опытах среднее время перевода отдельных слов при слуховом их восприятии с этой величиной, то можно сделать вывод о том, что время перевода отдельного слова практически совпадает со временем его прочтения на английском языке. Иначе говоря, сам процесс перевода происходит мгновенно.
Вторым моментом, выступившим в анализе полученных экспериментальных материалов, оказался факт неприменимости закона Хика к процессу актуализации лингвистических единиц. Уже говорилось выше, что этот закон перестает действовать при определенном количестве сигналов и реакций в эксперименте с реакцией выбора. Особенно разительно неприменимость закона Хика к анализу сложной реактивной деятельности человека проявляется в нашем эксперименте, в котором имеют место сотни и тысячи альтернатив. Если иметь в виду, что среднее время реакции при десяти сигналах составляет 0,62 с, то ока-жется, что основная масса значений времени речевых реакций при ог-ромном, практически бесконечном количестве альтернатив лежит ниже этой величины.
Объем словаря и количество возможных 'сигналов в ситуации про-веденного эксперимента таковы, что для объяснения полученных временных данных не может быть использована та модель последовательного представления реакций, их оценки и выбора их оптимальной, которая берет свое начало от Ф. Дондерса.
Невозможность в этих условиях последовательной деятельности по представлению различных вариантов и выбору из них наилучшего следует из сопоставления времени адекватных реакций на одиночные слова и речевые структуры со временем простой двигательной реакции на звуковой сигнал (0,14 с). Полученные в эксперименте длительности лишь в 3—6 раз превышают время простейшего психологического явления, в то время как количество хранимых в памяти реакций (адекватных русских ел ч) и число возможных сигналов (случайно подаваемых
английских слов) более тысячи. Полученные экспериментальные данные позволяют вместо последовательности перебора возможных реакций предложить иную модель актуализации прошлого опыта. Эта модель предполагает, что каждый входящий сигнал воздействует на всю совокупность прошлого опыта; в результате актуализируется та реакция (в нашем эксперименте то слово), которая адекватна пришедшему сигналу. Здесь возможна следующая аналогия: при воздействии определенным звуком на все струны рояля начинает вибрировать та струна, физические параметры которой соответствуют этому звуку. Именно такое взаимодействие в физике носит название резонансного взаимодействия. Следовательно, материалы проведенного эксперимента свидетельствуют в пользу волнового способа кодирования в психической деятельности.
Можно было бы привести много примеров резонансного управления, а следовательно, и волнового кодирования информационных процессов мозга. О реальности волновой психофизики говорят экспериментальные и теоретические материалы. Однако уже данных приведенного исследования достаточно для того, чтобы ощутить реальность волновых процессов, обеспечивающих регулирующую работу мозга.
Разумеется, самого факта резонанса в мозговых процессах еще недостаточно для доказательства голо графической природы образов и мыслей человека. Факты резонансных, основанных на волновом кодировании взаимодействий лишь в определенной мере повышают вероятность голографических моделей.
Прямым доказательством существования мозговых голограмм была бы прямая регистрация выбросов психофизических структур за пределы мозга. Большой методический интерес в этом отношении представляют так называемые пси-фото графин, сведения о которых время от времени появляются на страницах парапсихологических изданий (141, 158). Безусловно, подтверждающими эту гипотезу являются те факты засветки запечатанной фотографической пленки с помощью глаз, которые были зарегистрированы у известного экстрасенса и феномена Н. С. Кулагиной.
Все эти факты позволяют ставить вопрос о разработке такой методики, с помощью которой можно было бы осуществлять прямую регистрацию стоячих волн, выбрасываемых из глаз во время зрительного восприятия или представления различных объектов. При этом необходимо, чтобы такая регистрация была возможна не только у отдельных исключительных личностей, но и у обычных людей.
Разработка такого метода существенно продвинет вперед понимание работы мозга и прольет свет на многие парапсихологические явления. Если образ того или иного предмета, генерируемый мозгом человека, действительно окажется стоячей волной, своего рода голограммой, то к этому образу могут быть применены все те принципы, которые применяются к волновым структурам. Так, на образ как психофизи-40
|ческую категорию может быть распространен тот принцип нелокально-гсти, о котором мы говорили выше и согласно которому стоячие вол-Гны могут оказаться в любой точке пространства.
Реализуя принцип нелокальности в отношении психологических образов как голограмм, можно утверждать, что в каждой точке про-странства в латентном, скрытом виде существуют образы и мысли всех людей. Отсюда вытекает одна из гипотез, которая позволяет дать естественнонаучное объяснение биоинформационным контактам (телепатии). Разумеется, проблема регистрации образов как выброшенных, экстериоризованных за пределы мозга стоячих волн не является простой проблемой в методическом отношении. Исследователям еще придется решить целый ряд более мелких проблем, прежде чем будут созданы приборы, надежно регистрирующие мозговые голограммы. Но здесь научной психологии оказывает помощь совокупность материалов, полученных парапсихологами при изучении различных пси-феноменов, а также данные в психиатрии и психологии. Материалы эти при всей их исключительности свидетельствуют о реальности выдвигаемой ш ^холого-голо-графической гипотезы и о том, что поиски средств объективной регистрации образов у обычных испытуемых в конце концов увенчаются успехом.
В этом направлении сделаны очень важные наблюдения. Например, врач-психиатр Г. П. Крохалев проводил экспериментальные исследования галлюцинаций у больных людей и обнаружил, что зрительные галлюцинации могут быть объективно зарегистрированы на фото-и кинопленке (207, 208, 203). По мнению этого исследователя, глаз формирует в пространстве го л о графическое изображение образа, возникающего в мозге. Ученый считает, что при зрительных галлюцинациях происходит обратная передача информации от центра зрительного анализатора к периферии с проекцией зрительного образа из сетчатки глаз в пространство.
ГЛАВА 1. БИОИНФОРМАЦИОННЫЙ КОНТАКТ ЧЕЛОВЕК-РАСТЕНИЕ
Экспериментам, проводившимся в Советском Союзе и посвященным психолого-ботаническим дистанционным взаимодействиям человека и растения, предшествовали многочисленные фундаментальные исследования. К ним необходимо прежде всего отнести работы известного индийского ученого Дж. Чандра Боса, сделавшего крупнейшие открытия чувствительности и раздражимости растений и их связи с человеком. Существенен здесь также, цикл работ советских физиологов и биофизиков растений И. И. Гунара, В. Г. Карма-нова (316, 318) и ряда других, которые показали, что электрические импульсы и процессы, возникающие в растениях, имеют много общего с электрическими процессами, происходящими в организме животных и человека.
Чрезвычайно важным звеном с раскрытии механизмов биоинформационных взаимодействий являются исследования сибирского ученого В. П. Казначеева и его сотрудников (319). Эта группа исследователей, как известно, установила наличие информационной связи между клетками, помещенными в изолированные колбы. В этих экспериментах был зафиксирован факт как бы „сопереживания" между живыми клетками; отрицательное воздействие на одну клеточную культуру вызывало аналогичные изменения в клетках другой культуры.
В связи с этими экспериментальными результатами перестают казаться столь невероятными и сенсационными результаты американского исследователя К. Бакстера (27). Его результаты широко известны, они подробно описаны в книге П. Томпкинса и Кр. Берда „Тайная жизнь, растений" (28). Здесь целесообразно рассказать об экспериментах К. Бакстера лишь в самых общих чертах.
Все началось с того, что Бакстеру пришла довольно необычная мысль: поставить датчики, которые обычно ставят на человека при проведении опыта по детекции лжи, на комнатное растение. Ему захотелось выяснить, не возникнет ли у растения кожно-гальваническая реакция в тот момент, когда рядом будет умирать живое существо. Если бы такая реакция возникла, то криминалистика смогла бы получить новое мощное оружие: растения могли бы использоваться в качестве свидетелей преступлений. Эксперимент, в котором моделировалось наиболее тяжкое преступление - убийство, был организован следующим образом. Живая креветка располагалась на поверхности, закрепленной над сосудом с кипящей водой. Эта поверхность переворачивалась по сигналу датчика случайных чисел, т. е, в момент, не известный даже экспериментатору. Когда автомат срабатывал, креветка падала в кипящую воду и погибала, а на ленте прибора, предназначенного для регистрации КГР растения, появлялась отметка.
В этих экспериментах было зарегистрировано, что лист комнатного растения, находящегося вблизи умирающей креветки, реагирует на ее смерть тем самым кожно-гальваническим рефлексом, наличием которого характеризуются психоэнергетические процессы человека.
То ошеломляющее действие, которое оказали на весь мир, а отнюдь не только на научные круги эксперименты Бакстера, вполне понятно. Ведь растение представляет собой систему живых клеток, лишенных специализированной нервной регуляции. Что же касается креветки, то это — животное, обладающее пусть примитивной, но все же нервной системой. Эти живые организмы, стоящие на различных ступенях эволюционной лестницы, оказываются способными в определенном смысле „понимать" друг друга, „общаться" между собой на каком-то едином языке.
Еще большее впечатление произвел другой, также отмеченный К. Бакстером факт. Это факт контакта между растением и человеком. Из экспериментов американского криминалиста — а Бакстер был криминалистом-специалистом по детекции лжи - следовало, что те процессы, которые происходят в мозге человека и приводят к психоэнергетической реакции кожи, способны вызывать аналогичную реакцию у растения.
Совершенно естественно, что опыты Бакстера стали объектом пристального внимания со стороны многих исследователей в разных странах мира. Во многих лабораториях и университетах делались попытки воспроизведения биоинформационных контактов человека—растения.
Однако результаты проверочных экспериментов были неоднозначны. Некоторые лаборатории сообщали об удачном воспроизведении экспериментов Бакстера и подтверждали его основные результаты. Другие исследовательские коллективы сообщали о неподтверждении такого рода контактов. В ряде публикаций приводились сведения о том, что и сам Бакстер не всегда мог обеспечить стабильную повторяемость своих экспериментов: биоинформационный контакт то появлялся, то исчезал, хотя условия эксперимента всегда воспроизводились однозначно.
Создалась довольно сложная ситуация: с одной стороны, описанные Бакстером факты в ряде случаев подтверждались, а с другой — их воспроизводимость не была абсолютной. Эта ситуация требовала новой методики исследований, которая позволяла более глубоко и надежно
82
регистрировать воспроизводимость этой формы биоинформационных взаимодействий. Именно в такой неопределенной ситуации В. Н. Пушкин, В. М. Фетисов, Г. И. Ангушев начали свои поисковые психолого-ботанические исследования. Способ регистрации кожно-гальванической реакции в этих экспериментальных исследованиях был несколько отличен от записи КГР у Бакстера. Выше, при характеристике психоэнергетической функции кожи, указывалось, что существует два способа регистрации КГР — регистрация уменьшения кожного сопротивления (регистрация по Фере) и запись собственных импульсов кожи (регистрация по Тарханову). Если в экспериментах Бакстера использовался метод Фере, то в наших экспериментах был применен метод Тарханова: электрические реакции растения регистрировались с помощью четырех-канального энцефалографа.
Нужно отметить, что первые наши попытки зарегистрировать информационный контакт человека и растения были неудачными. Эти
9-09-2015, 17:52