Реферат
по анатомии ЦНС
на тему
Филогенез и онтогенез коры больших полушарий
Москва, 2010 г.
Оглавление
Введение
Общая информация о коре больших полушарий
Строение коры
Новая кора
Ассоциативные зоны и локализация полей
Филогенез коры
Онтогенез коры
Формирование коры в ходе внутриутробного развития ребенка
Первичные, вторичные и третичные поля коры
Заключение
Библиография
филогенез онтогенез кора полушарие
Введение
Кора головного мозга осуществляет тончайшее равновесие между организмом и внешней средой, регулирует и направляет физиологические процессы, протекающие внутри организма, обеспечивает его сложнейшее функциональное единство. В рамках данного реферата ставится цель посмотреть на процессы развития коры головного мозга в рамках большого периода филогенетического развития коры, а также индивидуального развития коры в рамках онтогенеза. Также ставится цель сравнить данные процессы с целью нахождения общих принципов и сходных процессов.
Общая информация о коре больших полушарий
Большие полушария – парные образования головного мозга. У человека они достигают ≈ 80% от общей массы мозга. Большие полушария осуществляют регуляцию высших нервных функций, лежащих в основе всех психических процессов человека, в то время как стволовая часть мозга обеспечивает низшие функции нервной системы, связанные с регуляцией деятельности внутренних органов.
Высшие функции обеспечиваются деятельностью особого отдела больших полушарий – коры головного мозга, которая несет главную ответственность за формирование условно-рефлекторных реакций. У человека по сравнению с животными кора одновременно ответственна и за согласование работы внутренних органов. Такое возрастание роли коры в регуляции всех функций в организме называется кортикализацией функций.
Кора выполняет следующие функции:
1. взаимодействие организма с внешней средой за счет безусловных и условных рефлексов.
2. осуществление высшей нервной деятельности (поведения) организма.
3. выполнение высших психических функций (мышления и сознания).
4. регуляция работы внутренних органов и обмена веществ в организме.
Кора больших полушарий представлена 12-18 миллиардами клеток, расположенных тонким слоем 3-4 мм на площади 2400 см2. 65-70 % этой площади находится в глубине борозд, а 30-35 % - на видимой поверхности полушарий. Кора состоит из нервных клеток, их отростков и нейроглинов, для которых характерно обилие межнейронных связей.
Строение коры
Кора покрывает поверхность больших полушарий с ее многочисленными бороздами и извилинами, за счет которых площадь коры значительно увеличивается. Различают ассоциативные зоны коры, а также сенсорную и моторную кору – области, в которых сосредоточены нейтроны, иннервирующие различные части тела.
Кора больших полушарий связана нервными путями со всеми нижележащими отделами центральной нервной системы, а через них — со всеми органами тела. С одной стороны, импульсы, поступающие с периферии, доходят до той или иной точки коры, с другой — кора посылает «распоряжения» в нижележащие отделы мозга, а оттуда — к различным органам.
По происхождению и структуре кора больших полушарий неоднородна. Большую часть коры у человека занимает новая кора неокортекс (neocortex), филогенетически наиболее молодая корковая формация. Филогенетически более ранние корковые структуры — древняя кора (paleocortex) и старая кора (archicortex) — занимают небольшую часть поверхности полушарий. Закладка новой коры образуется в латеральных частях плаща. Новая кора интенсивно развивается и оттесняет древнюю кору на основание полушарий, где она сохраняется в виде узкой полоски обонятельной коры и занимает 0,6% поверхности коры на вентральной поверхности полушарий, а старая кора отодвигается на медиальные поверхности полушарий, занимает 2, 2% поверхности коры и представлена гиппокампом и зубчатой извилиной. По происхождению и клеточному строению новая кора отличается от древней и старой коры. Переход от одной корковой формации к другой в клеточном строении происходит постепенно. Кора переходного типа называется межуточной корой , она занимает 1, 3% общей площади коры. Таким образом, большую часть поверхности коры (95, 6%) занимает новая кора.
Для древней коры характерно отсутствие послойного строения. В ней преобладают крупные нейроны, сгруппированные в клеточные островки. Старая кора имеет три клеточных слоя. Ключевой структурой старой коры является гиппокамп. Гиппокамп имеет обширные связи со многими другими структурами мозга. Он является центральной структурой лимбической системы мозга.
Функциональной единицей коры является вертикальная колонка взаимосвязанных нейронов. Все нейроны вертикальной колонки отвечают на одно и тоже афферентное раздражение одинаковой реакцией и совместно формируют эфферентный ответ. Распространение возбуждения в горизонтальном направлении (иррадиация) обеспечивается поперечными волокнами, идущими от одной вертикальной колонки к другой, а ограничивается – процессами торможения. Возникновение возбуждения в вертикальной колонке нейронов приводит к активности спинальные мотонейроны и к сокращению связанных с ними мышц.
Упорядоченное положение клеток в коре называется цитоархитектоникой, а их волокон – миелоархитектоникой.
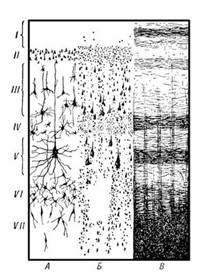
Рисунок 1. Схема цитоархитектонического строения новой коры головного мозга
На рисунке 1 представлена схема цитоархитектонического строения новой коры. В разделе А окрашены только тела клеток, Б окрашены отдельные клетки с отростками; В окрашены волокна. Слои коры: I — молекулярный слой; II — наружный зернистый слой; III — пирамидный слой; IV — внутренний зернистый слой; V — ганглионарный слой; VI — полиморфный слой.
Новая кора
Все области новой коры построены по единому принципу. Исходным типом является шестислойная кора (рис. 1). Слои представлены следующим образом:
Iслой — самый поверхностный, толщиной около 0,2 мм, называется молекулярным (lamina molecula ris). Он состоит из волокон апикальных дендритов и аксонов, поднимающихся от клеток нижних слоев, которые контактируют друг с другом. Нейронов в молекулярном слое незначительное количество. Это мелкие горизонтальные клетки и клетки-зерна. Все отростки клеток молекулярного слоя располагаются в пределах этого же слоя.
IIслой— наружныйзернистый(lamina granula ns exte rna).Толщина наружного зернистого слоя — 0,10 мм. Он состоит из мелких пирамидных и звездчатых нейронов. Аксоны этих нейронов оканчиваются на нейронах III, V и VI слоев.
III слой — пирамидный {lamina pyramidalis),толщиной около 1 мм, состоит из мелких и средних пирамидных клеток. Типичный пирамидный нейрон имеет форму треугольника, вершина которого направлена вверх. От вершины отходит апикальный дендрит, ветвящийся в вышележащих слоях. Аксон пирамидной клетки отходит от основания клетки и направляется вниз. Дендриты клеток III слоя направляются во второй слой. Аксоны клеток III слоя оканчиваются на клетках нижележащих слоев или образуют ассоциативные волокна.
IV слой — внутренний зернистый {lamina granula ns inte rnus).Он состоит из звездчатых клеток, имеющих короткие отростки, и малых пирамид. Дендриты клеток IV слоя уходят в молекулярный слой коры, а их коллатерали ветвятся в своем слое. Аксоны клеток IV слоя могут подниматься в вышележащие слои или уходить в белое вещество как ассоциативные волокна. Толщина IV слоя от 0,12 до 0,3 мм.
V слой — ганглионарный {lamina gangliona ris) — слой больших пирамид. Самые крупные клетки коры расположены именно в этом слое (гигантские пирамиды Беца передней центральной извилины). Их апикальные дендриты достигают молекулярного слоя, а базальные дендриты распределяются в своем слое. Аксоны клеток V слоя покидают кору и являются ассоциативными, комиссуральными или проекционными волокнами. Толщина V слоя достигает 0,5 мм.
VI слой коры — полиморфный {lamina multifo rmis).Содержит клетки разнообразной формы и размера, имеет толщину от 0,1 до 0,9 мм. Часть дендритов клеток этого слоя достигает молекулярного слоя, другие же остаются в пределах IV и V слоев. Аксоны клеток VI слоя могут подниматься к верхним слоям или уходить из коры в качестве коротких или длинных ассоциативных волокон.
Клетки одного слоя коры выполняют сходную функцию в обработке информации. I и IV слои являются местом ветвления ассоциативных и комиссуральных волокон, т.е. получают информацию от других корковых структур. III и IV слои являются входными, афферентными для проекционных полей, так как именно в этих слоях заканчиваются таламические волокна. V слой клеток выполняет эфферентную функцию, его аксоны несут информацию к нижележащим структурам мозга. VI слой также является выходным, но его аксоны кору не покидают, а являются ассоциативными.
Основным принципом функциональной организации коры является объединение нейронов в колонки. Колонка расположена перпендикулярно поверхности коры и охватывает все ее слои от поверхности к белому веществу. Связи между клетками одной колонки осуществляются по вертикали вдоль оси колонки. Боковые отростки клеток имеют небольшую длину. Связь между колонками соседних зон осуществляется через волокна, уходящие вглубь, а затем входящие в другую зону, т.е. короткие ассоциативные волокна. Функциональная организация коры в виде колонок обнаружена в соматосенсорной, зрительной, моторной и ассоциативной коре. Отдельные зоны коры имеют принципиально одинаковое клеточное строение, однако есть и отличия, особенно в структуре III, IV и V слоев, которые могут распадаться на несколько подслоев. Кроме этого, существенными цитоархитектоническими признаками являются плотность расположения и размеры клеток, наличие специфических типов нейронов, расположение и направление хода миелиновых волокон.
Цитоархитектонические особенности позволили разделить всю поверхность коры на 11 цитоархитектонических областей, включающих в себя 52 поля (по Бродману). Каждое цитоархитектоническое поле обозначено на картах мозга номером, который присваивался ему в порядке описания.
В первом слое коры волокна образуют полоску молекулярной пластинки. Во втором слое залегают тонкие волокна наружной зернистой пластинки. В составе четвертого слоя коры находится полоска внутренней зернистой пластинки (наружная полоска Байярже). В пятом слое содержатся волокна внутренней пирамидной пластинки (внутреняя полоска Байярже).
Основная информация в кору поступает по специфическим афферентным проводящим путям, заканчивающимся на клетках 3 и 4 слоев. Неспецифические пути от РФ заканчиваются в верхних слоях коры и регулируют ее функциональное состояние (возбуждение, торможение).
Звездчатые нейроны выполняют главным образом чувствительную (афферентную) функцию. Пирамидные и веретеновидные клетки – это преимущественно двигательные (эфферентные) нейроны.
Часть клеток коры принимают информацию от любых рецепторов организма – это полисенсорные нейроны, воспринимающие импульсы только от определенных рецепторов (зрительных, слуховых, тактильных и т.д.). Клетки нейроглии выполняют вспомогательные функции: трофическую, нейросекреторную, защитную, изолирующую.
Специализированные нейроны и другие клетки, входящие в состав вертикальных колонок, образуют отдельные участки коры, которые называются проекционными зонами (цитоархитектоническими полями)[1] . Эти функциональные зоны коры делятся на 3 группы:
– афферентные (чувственные);
– эфферентные (двигательные или моторные);
– ассоциативные (соединяют предыдущие зоны и обусловливают сложную работу мозга, лежащую в основе высшей психической деятельности).
Ассоциативные зоны и локализация полей
Каждый анализатор (например, зрения, обоняния, слуха и т.д.) имеет, по представлению И. П. Павлова, в коре головного мозга центральную часть (ядро), где осуществляется высший анализ и синтез, и широкую периферическую зону, в которой аналитические и синтетические процессы совершаются в элементарном виде. Между ядрами отдельных анализаторов разбросаны и перемешаны нервные элементы, принадлежащие различным анализаторам. Если ядро анализатора в силу каких-либо причин разрушено или выбыло из строя, его функцию перенимают периферические элементы того же анализатора. Современная физиология отвергает и узкий («абсолютный») локализационизм и принцип однородности, равноценности всех участков коры мозга. Локализация существует, но имеет «подвижный», «динамический» характер, о чем еще много лет назад говорил И.П. Павлов.
Нервные образования, которые мы привыкли называть «центрами», не ограничиваются корой головного мозга. Они включают и подкорковые структуры, значение которых необычайно велико.
Следует помнить, что любой центр коры головного мозга теснейшим образом связан со всеми другими отделами центральной нервной системы. В этом объединении, или, как говорят, интеграции, и заключается ведущая роль коры мозга в организме. Представление о единых корковых центрах, полностью обеспечивающих какую-либо определенную функцию, является в настоящее время пройденным этапом в физиологии.
К тому же кора головного мозга отличается необычайной пластичностью, и одни отделы ее легко перенимают функции других, компенсируя расстройство их деятельности, вызванное различными причинами. Наиболее важная задача современной науки выявить анатомическую основу физиологических процессов и одновременно установить связи и взаимосвязи между всеми явлениями, наблюдаемыми в головном мозгу. Исследования, проведенные различными авторами, как отечественными, так и зарубежными, показали, что в центральной извилине мозга, расположенной спереди от центральной борозды, находится специальная двигательная область. Раздражение ее электрическим током вызывает сокращение определенных мышц противоположной стороны тела. Напротив, удаление этой области хирургическим путем ведет к расстройству координированных движений, шаткости походки, ослаблению мышц. У человека ранение двигательной области сопровождается обычно параличами и другими тяжелыми нарушениями деятельности организма. С помощью метода условных рефлексов удалось показать, что так называемые двигательные центры содержат чувствительные клетки, к которым приходят периферические раздражения от двигательного аппарата (костей, суставов, мышц). Эта область является мозговым концом двигательного анализатора в такой же степени, как затылочная — мозговым концом зрительного анализатора, височная — слухового анализатора и т.д. В двигательной области имеются как чувствительные клетки, расположенные в верхних слоях коры, так и двигательные, сосредоточенные в ее нижних слоях. Импульсы от рецепторов двигательного аппарата поступают в чувствительные клетки передней мозговой извилины, а отсюда уже передаются двигательным клеткам головного и спинного мозга.
У человека ассоциативные зоны достигают наибольшего развития. Локализация функций в коре головного мозга относительна – здесь нельзя провести каких-либо четких границ, поэтому мозг обладает высокой пластичностью, приспосабливаемостью к повреждениям. Тем не менее, морфологическая и функциональная неоднородность коры позволила выделить в ней 52 цитоархитектонических поля (К. Бродман), а среди них – центры зрения, слуха, осязания и др. Все они связаны между собой волокнами проводящих путей белого вещества, которые делятся на 3 типа:
1. ассоциативные (связывают зоны коры в пределах одного полушария),
2. комиссуральные (связывают симметричные зоны коры двух полушарий через мозолистое тело),
3. проекционные (связывают кору и подкорку с периферическими органами, бывают чувствительные и двигательные).
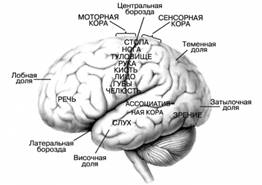
Рисунок 2. Значение важнейших зон коры головного мозга
1. Чувствительная зона коры (в постцентральной извилине) воспринимает импульсы от тактильных, температурных и болевых рецепторов кожи, а также от проприорецепторов противоположной половины тела.
2. Двигательная зона коры (в предцентральной извилине) содержит в 5 слое коры пирамидные клетки Беца, от которых идут импульсы произвольных движений к скелетным мышцам противоположной половины тела.
3. Премоторная зона (в основании средней лобной извилины) обеспечивает сочетанный поворот головы и глаз в противоположную сторону.
4. Праксическая зона (в надкраевой извилине) обеспечивает сложные целенаправленные движения практической деятельности и профессиональных двигательных навыков. Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
5. Центр проприоцептивного гнозиса (в верхней теменной дольке) обеспечивает восприятие импульсов проприорецепторов, контролирует ощущения тела и его частей как целостного образования.
6. Центр чтения (в верхней теменной дольке, вблизи затылочной доли) контролирует восприятие написанного текста.
7. Слуховая зона коры (в верхней височной извилине) воспринимает информацию от рецепторов органа слуха.
8. Слуховой центр речи, центр Вернике (в основании верхней височной извилины). Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
9. Слуховой центр пения (в верхней височной извилине). Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
10. Двигательный центр устной речи, центр Брока (в основании нижней лобной извилины) контролирует произвольные сокращения мышц, участвующих в речеобразовании. Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
11. Двигательный центр письменной речи (в основании средней лобной извилины) обеспечивает произвольные движения, связанные с написанием букв и других знаков. Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
12. Стереогностическая зона (в угловой извилине) контролирует узнавание предметов наощупь (стереогноз).
13. Зрительная зона коры (в затылочной доле) воспринимает информацию от рецепторов органа зрения.
14. Зрительный центр речи (в угловой извилине) контролирует движение губ и мимику говорящего оппонента, тесно связан с другими сенсорными и моторными речевыми центрами. Речь и сознание – это филогенетические наиболее молодые функции мозга, поэтому речевые центры имеют большое число рассеянных элементов и наименее локализованы. Речевые и мыслительные функции выполняются при участии всей коры. Речевые центры у человека сформировались на основе трудовой деятельности, поэтому они асимметричные, непарные и связаны с рабочей рукой.
Позади центральной борозды расположена чувствительная область коры. В ней заканчивается путь, начавшийся в рецепторах кожи и внутренних органов. Здесь расположен его конечный этап. Каждое полушарие мозга связано в основном с противоположной половиной тела. Однако существуют связи полушария и с одноименной половиной тела.
Филогенез коры
Кора — продукт длительного эволюционного развития, в процессе которого сначала появляется древняя кора, возникающая в связи с развитием обонятельного анализатора у рыб. С выходом животных из воды на сушу начинает интенсивно развиваться т. н. плащевидная, полностью обособленная от подкорки часть коры, которая состоит из старой и новой коры. Становление этих структур в процессе приспособления к сложным и разнообразным условиям наземного существования связано (совершенствованием и взаимодействием различных воспринимающих и двигательных систем. У земноводных кора представлена древней и зачатком старой коры, у пресмыкающихся хорошо развиты древняя и старая кора и появляется зачаток новой коры. Наибольшего развития новая кора достигает у млекопитающих, а среди них у приматов (обезьяны и человек), хоботных (слоны) и китообразных (дельфины, киты). В связи с неравномерностью роста отдельных структур новой коры её поверхность становится складчатой, покрываясь бороздами и извилинами. Совершенствование коры конечного мозга у млекопитающих неразрывно связано с эволюцией всех отделов центральной нервной системы. Этот процесс сопровождается интенсивным ростом прямых и обратных связей, соединяющих корковые и подкорковые структуры. Т. о., на более высоких этапах эволюции функции подкорковых образований начинают контролироваться корковыми структурами. Данное явление получило название кортиколизации функций. В результате кортиколизации ствол мозга образует с корковыми
8-09-2015, 19:09