Авидин-биотиновая реакция в иммуноанализе
Введение
В последние годы в качестве альтернативы радионуклидам в иммуноанализе нашли широкое применение неизотопные метки, в том числе ферменты, хемилюминесцентные соединения, флуорофоры и коллоидные частицы. Применение неизотопных маркеров затрудняется необходимостью синтезировать, очищать и охарактеризовывать белковые конъюгаты в случае каждого определяемого вещества. Ранее система авидин – биотин нашла широкое применение при локализации, обнаружении и очистке белков. Оказалось, что эта система не менее полезна и в иммуноанализе.
В работе детально рассмотрены два подхода, с помощью которых авидин-биотиновая реакция может быть использована в двухцентровом иммуноанализе. В обоих подходах применяли препараты иммобилизованных и биотинилированных антител. Эти препараты отбирали по принципу распознавания различных эпитопов антигена. В первом подходе сначала иммобилизованные антитела вводили в реакцию с исследуемым образцом и биотинилированными антителами. Затем добавляли конъюгат авидина или стрептавидина с ферментом или с хемилюминесцентным маркером. Во втором подходе вместо белкового конъюгата добавляли свободный авидин или стрептавидин и затем добавляли биотинилированный фермент. В обоих случаях определение осуществляли спектрофотометрически или по интенсивности люминесценции после добавления соответствующих субстратов. В качестве биотинилированных ферментов-маркеров применяли пероксидазу хрена, АВ+ -зависимые ферменты, АТР-зависимые ферменты, пенициллиназу и щелочную фосфатазу. Рассмотрим некоторые результаты, полученные с помощью этих двух подходов.
Основные принципы
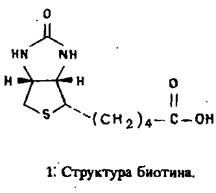
Уже давно известно, что для обеспечения сильного взаимодействия биотина с гликопротеином яичного белка авидином требуется только уреиднос кольцо витамина. Следовательно, карбоксильную группу остатка валериановой кислоты в биотине можно модифицировать и таким образом получать активные биотинилированные производные. В частности, биотин, связанный с макромолекулой, сохраняет способность связываться с активным центром авидина. Поскольку молекула авидина состоит из четырех идентичных субъединиц, то с ней могут быть одновременно связаны остатки биотина двух различных биотинилированных белков. Эти свойства авидии-биотинового комплекса широко используются для решения различных задач, в том числе 1) обнаружения, локализации и очистки белков, углеводов и нуклеиновых кислот; 2) разработки методов двухцентрового иммунометрического анализа; 3) картирования эпигонов в гибридомной технологии.
Практическое применение авидин-биотиновой системы ограничивается высокой основностью авидина и наличием в его молекуле углеводных остатков, что в свою очередь обусловливает высокий уровень неспецифического связывания. Более 20 лет назад в бактериях Streptomycesavidiniiбыл обнаружен другой биотинсвязывающий белок, названный стрептавидином. Подобно авидину яичного белка, стрептавидин также образует очень прочный и специфический нековалент-ный комплекс с биотином и состоит из четырех идентичных субъединиц. Каждая субъединица содержит один биотинсвязываюший центр. В отличие от авидина стрептавидин негликозилирован и имеет нейтральную ph. Авидин и стрептавидин сильно различаются по аминокислотному составу, хотя оба богаты остатками триптофана. Некоторые свойства этих белков приведены в табл. 1.
Таблица 1. Некоторые характеристики авидина и стрептавидина
| Характеристики | Авидин | Стрептавиднн |
| tМолекулярная масса | 67 000 | 60 000 |
| Мол. масса субъединицы | 15 600 | 14 600 . |
| Kd (авидин-биотиновый комплекс), М | -10-15 | -10-15 |
| Еш (1 мг/мл) | 1,54 | 3,4 |
| Связывание биотина на субъединицу | 1 | 1 |
| Олигосахариды на субъединицу | 1 | 0 |
| Манноэа на субъединнцу | 4-5 | 0 |
| GlcNAc* ва субъединицу | 3 | 0 |
| Иэоэлектрическая точка, pi | >10 | <7 |
| Остатки аминокислот на субъеднницу | ||
| Триптофавы | 4 | 8 |
| Тироэины | 1 | 6 |
| Лиэины | 9 | 4 |
| Аргинины | 8 | 4 |
| Аспарагин + аспарагиновая кислота | 15 | 12 |
| Глутамин + глутаминовая кислота | 10 | 9 |
| Амиды | 16 | - |
Стрептавидин можно использовать вместо авидина или в дополнение к нему в авидин-биотиновых системах. Он коммерчески доступен, и, подобно авидину, его можно очищать с помощью аффинной хроматографии на иминобиотиновой колонке. Поскольку бактериальный белок имеет нейтральную изоэлектрическую точку и не содержит углеводных остатков, то для него нехарактерны неспецифические взаимодействия, свойственные авидину яичного белка. Хотя неспецифическое связывание авидина можно свести к минимуму с помощью химических или каких-либо других методов, стрептавиднн и его конъюгаты являются наилучшим решением этой проблемы.
В лаборатории для разработки различных методов иммуноанализа применяются имеющиеся в продаже конъюгаты авидина с ферментами, а также синтезированные препараты биотинилированных белков и стрептавидина с хемилюминесцентной меткой.
Получение биотинилированных белков
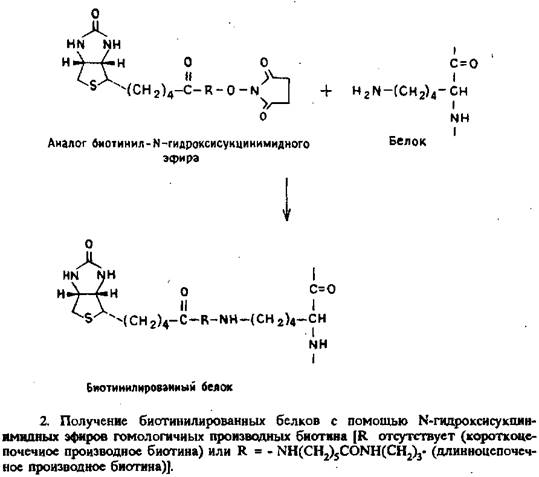
Антитела и ферменты биотинилировали по ранее описанной методике с использованием N-гидроксисукцинимидных эфиров производных биотина с различной длиной алкильной цепи. Структура реагентов и схема реакции представлены на рис. 2. Качество полученных препаратов проверяли с помощью ELISAили двухцентрового иммунометрического анализа.
Получение белков с хемилюминесцентной меткой
Стрептавиднн или очищенные фракции IgG моноклональных антител обрабатывали сукцинимидными производными аминобутилэтилизолюминола. Полученные конъюгаты очищали гельфильтрацией на сефадексе G-25 и хранили при -20°С.
Методы анализа
При разработке методов двухцентрового иммунометрического анализа белков применяли меченые конъюгаты авидина или стрептавидина либо стрептавидин как связующее звено между биотинилированными антителами и биотинилированным ферментом. Величину сигнала в ферментативной реакции как в том, так и в другом случае определяли спектрофотометрически или по интенсивности люминесценции в зависимости от типа используемой метки.
Авидин и стрептавидин в качестве меченых зондов
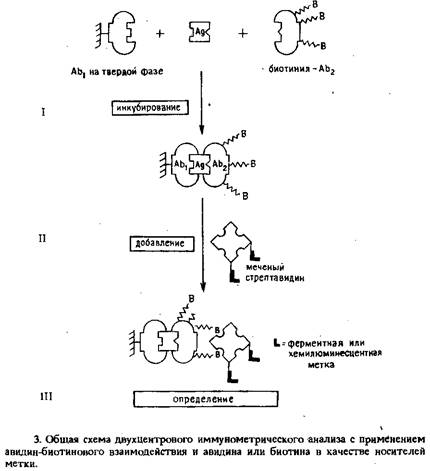
Принципиальная схема этого метода представлена на рис. 3.Исследуемый образец инкубировали с высокоаффинными моноклональными антителами, ковалентно связанными с полистирольными шариками диаметром 6,5 мм. Затем в систему вводили биотинилированные антитела к другим эпитопам антигена. По завершении иммунологической реакции добавляли авидин, меченный ферментом, или содержащий хемилюминесцентную метку стрептавидин. В зависимости от используемого маркера измерение осуществляли спектрофотометрически или по интенсивности люминесценции.
Определение с помощью спектрофотометрии
Для изучения влияния длины цепи между молекулой биотина и моноклональным антителом на чувствительность анализа применяли систему, в которой избыток моноклональных аити-hGH антител был ковалентно связан с полистирольными гранулами. Гранулы инкубировали со стандартными препаратами или с пробами и с двумя различными препаратами биотинилированных моноклональных hGH-антител, различающимися длиной цепи, соединяющей молекулу биотина и антитела. По завершении реакции антиген – антитело в систему добавляли конъюгатавидин – щелочная фосфатаза и смесь инкубировали в течение 30 мин. Растворы упаривали в токе воздуха, добавляли субстрат м-нитрофенилфосфат и смесь инкубировали 45 мин при 37°С.
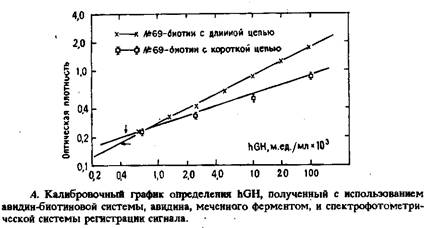
Образовавшийся продукт ферментативной реакции определяли по поглощению при 405 нм. Экспериментальные данные представляли в виде графика зависимости логарифма оптической плотности от логарифма концентрации hGH. Полученные данные свидетельствуют, что более крутой наклон наблюдается в случае антител клона №69, биотинилированных с помощью N-гидроксисукцинимидного эфира с длинной цепью CONH3 -). Чувствительность анализа, достигаемая с длинноцепочечными биотинилированными антителами клона №69, сравнима с чувствительностью определения гормона с помощью обычного конкурентного РИА.
Определение путем измерения интенсивности люминесценции
Кроме спектрофотометрии для определения связанных биотинилированных антител мы также использовали измерение интенсивности люминесценции. На этом рисунке символом L обозначен хемилюминесцентный маркер, а моноклональные антитела к hGHбиотинилировали через длинную цепочку CONH3-). В данном методе моноклональные антитела, связанные с полистирольными гранулами, инкубировали со стандартами или пробами и с длинноцепочечными производными биотинилированных антител анти-hGHпри комнатной температуре.
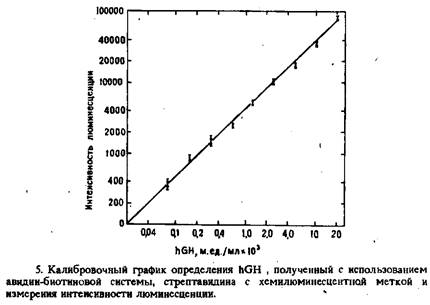
По завершении иммунологической реакции добавляли стрептавидин, несущий хемилюминесцентную метку. Через 30 мин гранулы промывали. Интенсивность свечения связанной метки регистрировали при щелочном рН с использованием системы микропероксидаза – пероксид водорода. Калибровочный график зависимости логарифма интенсивности хемилюминесценции от логарифма концентрации hGH представлен на рис. 5. Предел обнаружения гормона с помощью данного метода составляет 3-10-5 м.ед. hGH/л или 1,5 пг в пробирке, а рабочий диапазон – от 4–10-5 до 10-1 м.ед. hGH/л. Коэффициент вариации анализа внутри серии равен 5,5%.
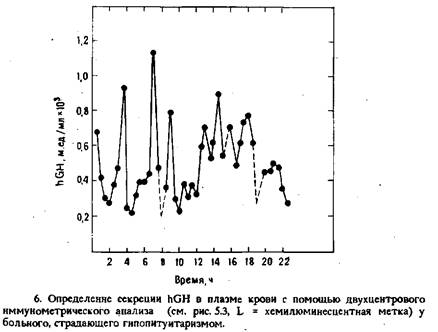
Нормальный уровень hGH и его динамические изменения при искусственной стимуляции у здоровых пациентов определяли с помощью обычного РИА и двухцентрового иммунометрического анализа. Коэффициент корреляции обоих методов составил 0,96. Кроме того, метод ИХМА позволил определить уровень базальной секреции и слабые изменения концентрации hGH в плазме крови у больных гипопитуитаризмом в течение циркадного цикла, что невозможно сделать с помощью обычного РИА. Полученные данные показывают, что люминесцентный ИХМА в 10 раз более чувствителен, чем существующие радио- и иммуноферментные методы определения hGH.
Стрептавидин и биотинилированные ферменты какзонды
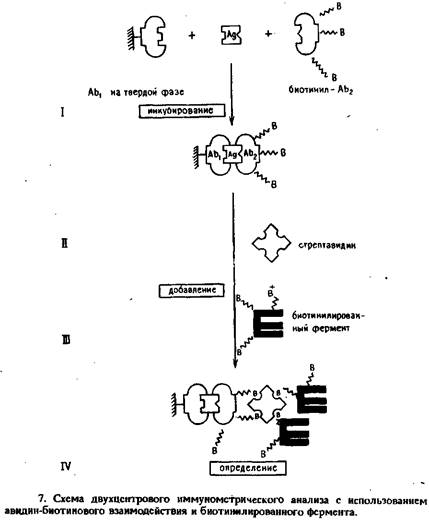
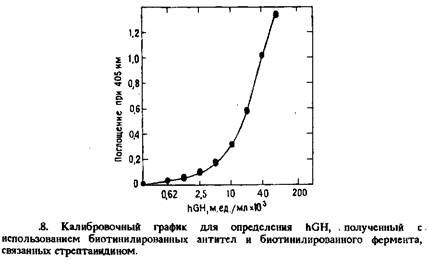
Принципиальная последовательность операций в этом методе представлена на рис 7. Связанные с носителем и биотинилированные моноклональные антитела инкубировали с пробой или стандартами. По завершении иммунологической реакции последовательно добавляли или стрептавидин и биотинилированную щелочную фосфатазу, или ацетилированный авидин и биотинилированную глюкозо-6-фосфатдегидрогеназу. В зависимости от фермента величину сигнала регистрировали спектрофотометрически или по интенсивности люминесценции. Примером такого подхода может служить определение hGHс помощью иммобилизованных на полистирольных гранулах моноклональных антител анти-hGHи модифицированных длинноцепочечным производным биотина антител анти-hGH клона №69. Гранулы отмывали и последовательно инкубировали при комнатной температуре со стрептавидином и длинноцепочечным производным биотинилированной щелочной фосфатазы. После промывки добавляли раствор j-нитрофенилфрсфата. Образование продукта регистрировали по поглощению при 405 нм. Калибровочный график для определения hGH, полученный с помощью биотинилированной щелочной фосфатазы и спектрофотометрического определения продукта ферментативной реакции, представлен на рис. 8.
Чувствительность анализа сравнима с чувствительностью методов типа РИА. Коэффициент вариации внутри серии при анализе смешанной плазмы с концентрацией hGH 7.6–10"3 м.ед./л равен 6,5%, а коэффициент вариации между сериями при анализе того же пула – 7,4%.
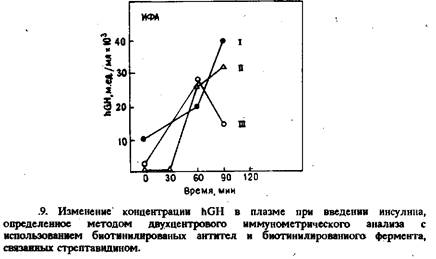
На рис. 9 приведены результаты анализа образцов плазмы, взятых у пациентов, подвергнутых тесту «на стимуляцию секреции гормона роста. Полученные данные свидетельствуют, что с помощью этого метода можно регистрировать и количественно определять реакцию в тестах по стимуляции и подавлению синтеза hGH, проводимых для оценки резервов гормона в гипофизе. Все применявшиеся в этом методе реагенты устойчивы в течение по меньшей мере двух лет. Диапазон калибровочной кривой позволяет определять в одном анализе высокие и низкие концентрации hGH.
Заключение
Сравнительные данные для двух методов двухцентрового иммунометрического анализа hGH приведены в табл. 2. Чувствительность использованного для сравнения метода РИА составляла 0,6–10-3 м.ед./л, а рабочий диапазон – 10-3 м.ед./л.
8-09-2015, 19:53