МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ УКРАИНЫ
ТАВРИЧЕСКИЙ НАЦИОНАЛЬНЫЙ УНИВЕРСИТЕТ
им. В.И.ВЕРНАДСКОГО
КЕРЧЕНСКИЙ ЭКОНОМИКО-ГУМАНИТАРНЫЙ ИНСТИТУТ
Факультет физической культуры и спорта
Филиал кафедры физической реабилитации
Бобер Степан Васильевич
ГИПОТАЛАМО – ГИПОФЗАРНАЯ РЕГУЛЯЦИЯ ФУНКЦИЙ ОРГАНИЗМА В ОНТОГЕНЕЗЕ
Курсовая работа
Дисциплина: «Физиология человека»
Специальность: 6.010200 «Физическая реабилитация»
Курс 2, группа 2005-Ф, дневная форма обучения
Регистрационный номер _____ Курсовая работа защищена
Курсовая работа с оценкой ______________
к защите допускается «___»_____________ 200 г
«___»____________ 200 г
Научный руководитель: Научный руководитель:
Козлова Г.В. Козлова Г.В.
_________________ _____________________
Керчь, 2007
Реферат
Бобер. С.В. «Гипоталамо – гипофизарная регуляция функций организма в онтогенезе». Работа изложена на 48 страницах машинописного текста, содержит 3 раздела, содержит 3 рисунка, 3 таблицы. Список использованных работ включает 26 источников.
Цель работы – показать значение для организма гипоталамо – гипофизарной регуляции, описать основные процессы, происходящие в нейрогуморальном аппарате и их влияние на основные гомеостатические процессы, и показать их воздействие на поведение, и на общее состояние организма в целом в процессе онтогенеза
Для решения поставленных задач использовался метод литературного обзора
ГИПОТАЛАМУС, ГИПОФИЗ, ЛИБЕРИНЫ, СТАТИНЫ, ПАРАКРИНЫ, ГОМЕОСТАЗ, ГОРМОН, ТУРЕЦКОЕ СЕДЛО, СИНАПС, ВИТАЛЬНЫЕ ЦЕНТРЫ
СОДЕРЖАНИЕ
Стр.
ВВЕДЕНИЕ……………………………………………………………………4
РАЗДЕЛ 1.МОРФО-ФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ ГИПОЛАМО – ГИПОФИЗАРНОГО АППАРАТА ………....…6
1.1.Функциональная анатомия гипоталамуса………………..........6
1.2.Строение и функции гипофиза, гормоны гипофиза…………..9
1.3.Взаимодеиствие и взаимоотношение гипоталамуса и гипофиза …………………………………………………………………………..……..18
РАЗДЕЛ 2.МЕТОДЫ ИССЛЕДОВАНИЯ ГИПОТАЛАМО – ГИПОФИЗАРНОЙСИСТЕМЫ………………………....26
2.1.Методы исследования желез внутренней секреции……….26
2.2.Иследование гипоталамуса в эксперименте………………..29
РАЗДЕЛ 3. РЕГУЛЯЦИЯ ГИПОТАЛАМО – ГИПОФИЗАРНОЙ СИСТЕМЫ ………..………………….……………32
3.1.Прямые и обратные связи в регуляции синтеза гормонов…. 32
3.2.Центральная регуляция гипоталамо – гипофизарной системы…………………………………….. 39
3.3.Гипоталамус и сердечно –сосудистая система……………….39
3.4.Гипоталамо – гипофизарный аппарат и поведение ………….41
ВЫВОДЫ……………………………………………………………………...44
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ………………………....48
ВВЕДЕНИЕ
Актуальность данной работы состоит в том, что функция и деятельность гипоталамо - гипофизарного аппарата неразрывно связана с основными гомеостатическими процессами и функционированием всех внутренних органов и правильное понятие о процессах происходящих в этой системе является необходимой для правильного понимания гомеостатических процессов происходящих в организме человека. Гипоталамо-гипофизарный аппарат – важнейшее образование в базальной части головного мозга, обладающее интеграционной функцией, направленной на обеспечение жизнедеятельности всего организма и поддержание основных гомеостатических процессов и констант. Гипоталамус, как первая составляющая этого аппарата представляет собой главный нервный центр, отвечающий за «прием» и анализ информации получаемой как от нервной системы (прежде всего афферентные пути приходящие в таламус) так и от основных жидких сред организма (прежде всего кровь и ликвор). Филогенетически это древний отдел головного мозга, поэтому у наземных позвоночных млекопитающих его строение относительно одинаково в отличие от организации таких, более молодых структур, как новая кора и лимбическая система. Последнее обстоятельство полностью объясняет тот факт, что при разрушении корковых структур, но с сохраненным гипоталамусом можно достаточно легко сохранить жизнь организму, в то же время для поддержания жизнедеятельности животного с удаленным гипоталамусом требуются особые интенсивные меры, так как у такого животного уничтожены основные гомеостатические механизмы.
Гипофиз, вторая составляющая, условно может быть отнесен к исполнительному отделу выше указанного аппарата. Его главная задача гипофиза синтез целого спектра биологически активных веществ – гормонов, «мишенями» которых в большинстве случаев являются железы внутренней секреции, или эндокринные.
Цель работы – показать значение для организма гипоталамо – гипофизарной регуляции, описать основные процессы, происходящие в нейрогуморальном аппарате и их влияние на основные гомеостатические процессы, и показать их воздействие на поведение, и на общее состояние организма в целом в процессе онтогенеза. Для решения поставленных задач использовался метод литературного обзора
В работе были поставлены следующие задачи:
1. Изучить на основе анализа научно – медицинской литературы, анатомо-физиологические особенности системы
2. Перечислить основные методы изучения деятельности желез внутренней секреции в современной науке, осветить наиболее распространенные.
3.Осветить актуальность правильного функционирования данной системы в процессе жизнедеятельности и развития организма, дать краткую характеристику основных процессов протекающих в эндокринной системе под воздействием гипоталамо – гипофизарного аппарата, описать влияние на гипоталамо – гипофизарную систему выше лежащих структур мозга, а также влияние гипоталамо – гипофизарной системы на сердечнососудистую систему и поведение.
РАЗДЕЛ 1 МОРФО – ФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ ГИПОТАЛАМО - ГИПОФИЗАРНОГО АППАРАТА
1.1. Функциональная анатомия гипоталамуса
Расположение и строение гипоталамуса. Гипоталамус представляет собой небольшой отдел головного мозга, массой около 5,0 г. Он не имеет четких границ, поэтому его стоит рассматривать как часть сети нейронов, протягивающейся от среднего мозга через гипоталамус к глубинным отделам переднего мозга, тесно связанным с филогенетически древней обонятельной системой. Гипоталамус является центральным отделом промежуточного мозга: он лежит ниже (вентральнее) таламуса, образуя нижнюю половину стенки третьего желудочка. Нижней границей гипоталамуса служит средний мозг, а верхней - конечная пластинка, передняя спайка и зрительный перекрест. Латеральнее гипоталамуса расположены зрительный тракт, внутренняя капсула и субталамические структуры.
В поперечном направлении гипоталамус можно разделить на три зоны -перивентрикулярную, медиальную и латеральную. (Рис. № 1) Перивентрикулярная зона представляет собой тонкую полоску, прилежащую к третьему желудочку. 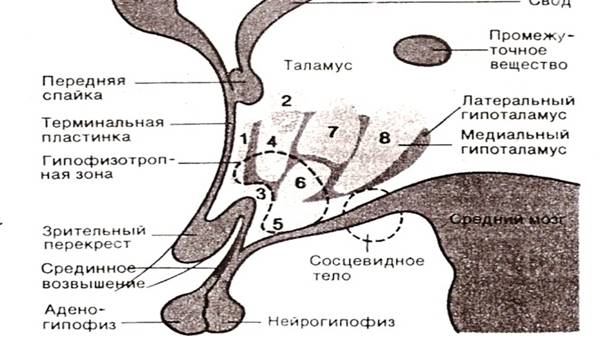
Рис.1.1. Ядерные зоны гипоталамуса на схеме сагиттальный разреза через третий желудочек.1- преоптическое ядро (приоптическая область);2 – паравентрикулярное ядро;3 –супраоптическое ядро;4 – преоптическое ядро;5 – инфундибулярное ядро;6 – вентромедиальное ядро;7 –дорсомедиальное ядро;8 – заднее ядро(задняя область)(по Benninahoff – Goertler.LehrbruchderMenschen,Vol. 3,UrbanandSchwarzenberg, 1977)
В медиальной зоне различают несколько ядерных областей, расположенных в передне-заднем направлении. Преоптическая область филогенетически относится к переднему мозгу, однако ее обычно относят к гипоталамусу.
От вентромедиальной области гипоталамуса начинается ножка гипофиза, соединяющаяся с адено - и нейрогипофизом. Передняя часть этой ножки носит название срединного возвышения. В срединном возвышении оканчиваются отростки многих нейронов преоптической и передней областей гипоталамуса, а также вентромедиального и инфундибулярного ядер. Здесь из этих отростков высвобождаются нейрогормоны поступающие через систему портальных сосудов к передней доле гипофиза (аденогипофизу). Совокупность ядерных зон, в которых содержатся гормон-продуцирующие нейроны, носит название гипофизотропной области (по Р Шмидт,Г. Тевс 1996. )
Нейрогормоны – это белковые, или пептидные биологически активные вещества, относящиеся к гормонам и секретируемые в кровеносное русло нервными клетками. Подробней о них будет сказано ниже.
Отростки нейронов супраоптического и паравентрикулярного ядер идут к задней доле гипофиза (нейрогипофиз). Эти нейроны регулируют образование и высвобождение окситоцина и вазопрессина. Конкретные функции отдельных ядер гипоталамуса, за исключением супраоптического и паравентрикулярного ядер, до настоящего времени окончательно не изучены.
В латеральном гипоталамусе не существует отдельных ядерных областей. Нейроны этой зоны располагаются диффузно вокруг медиального пучка переднего мозга, идущего в рострально-каудальном направлении от латеральных образований основания лимбической системы к передним центрам промежуточного мозга. Этот пучок состоит из длинных и коротких восходящих и нисходящих волокон
Организация афферентных и эфферентных связей гипоталамуса свидетельствует о том, что он служит важным интегративным центром для соматических, вегетативных и эндокринных функций.
Медиальный гипоталамус обладает двусторонними связями с латеральным гипоталамусом и, кроме того, непосредственно получает ряд сигналов из остальных отделов головного мозга. В медиальной области гипоталамуса существуют особые нейроны, воспринимающие важные параметры крови и спинномозговой жидкости. Иными словами, эти нейроны следят за состоянием внутренней среды организма. Они могут воспринимать, например, температуру крови, солевой состав плазмы или содержание гормонов в крови. (Сапин М.Р. 2005 ).
Здесь уместно дать характеристику системы кровоснабжения гипоталамуса. Большое значение в регуляции функций передней доли гипофиза имеют особенности ее кровоснабжения, а именно то, что кровь, оттекающая от капилляров гипоталамической области, поступает в так называемые портальные сосуды гипофиза и омывает его клетки. В гипоталамической области вокруг этих капилляров существует нервная сеть, состоящая из отростков нервных клеток, формирующих на капиллярах своеобразные нейрокапиллярные синапсы. Через эти образования продукты нейросекреции клеток гипоталамуса поступают в кровь и с ее током, переносятся к клеткам передней доли гипофиза, изменяя их функции.
Нейронам гипоталамуса, продуцирующим гормоны, присущи функции одновременно секреторных и нервных клеток. Это находит свое выражение в том, что в процессе секреции гормонов нервными клетками в них возникают потенциалы действия, аналогичные наблюдающимся при возникновении и распространении процесса возбуждения. Генерированием подобных потенциалов действия секреция железистых клеток никогда не сопровождается.
С общепринятой точки зрения считается, что посредством нервных механизмов медиальная область гипоталамуса управляет деятельностью нейрогипофиза, а посредством гормональных -аденогипофиза. Таким образом, эта область служит промежуточным звеном между нервной и эндокринной системами, представляя собой «нейроэндокринный интерфейс».
Латеральный гипоталамус образует двусторонние связи с верхними отделами ствола мозга, центральным серым веществом среднего мозга и с лимбической областью среднего мозга и с лимбической системой. Чувствительные сигналы от поверхности тела и внутренних органов поступают в гипоталамус по восходящим спинобульборетикулярным путям. Эти пути идут в гипоталамус либо через таламус, либо через лимбическую область среднего мозга. Остальные афферентные сигналы поступают в гипоталамус по полисинаптическим путям, которые пока еще не все идентифицированы. Эфферентные связи гипоталамуса с вегетативными и соматическими ядрами ствола мозга и спинного мозга образованы полисинаптическими путями, идущими в составе ретикулярной формации.( В. Яковлев 2006).
1.2. Строение и функции гипофиза, гормоны гипофиза.
Гипофиз состоит из трех долей - передней, промежуточной и задней, каждая из которых является железой внутренней секреции. Заднюю долю, богато снабженную разветвлениями нервных волокон, связывающих ее с гипоталамусом, часто называют нейрогипофизом, а переднюю чисто железистую долю - аденогипофизом.
Передняя доля . или аденогипофиз, состоит из главных или хромофобных клеток (55-60% всех клеток) и хромофильных: ацидофильных (30-35%) и базофильных (51%). Хромофобные клетки, по-видимому, гормонов не продуцируют и являются предшественниками хромофильных клеток. Ацидофильные клетки продуцируют соматотропный гормон и пролактин. Все гормоны передней доли являются белковыми веществами. Базофильные клетки продуцируют адренокортикотропный, тиреотропный и гонадотропный (фолликулостимулирующий и лютеинизирующий) гормоны. (по Р Шмидт,Г. Тевс 1996. )
Соматотропный гормон (гормон роста, соматотропин) стимулирует синтез белка в органах и тканях и рост молодых животных.
у соматотропного гормона хорошо выражена видовая специфичность. Препараты, полученные из гипофиза быка и свиньи, мало влияют или совсем не влияют на рост обезьяны и человека.
Соматотропин низших обезьян малоэффективен у человека, но гормон роста человека и высших обезьян ускоряет рост низших обезьян. Сделан вывод, что соматотропный гормон действует вниз и не действует вверх по эволюционной лестнице. Соматотропин повышает биосинтез рибонуклеиновой. кислоты - необходимого звена синтеза белков. Он усиливает транспорт аминокислот из крови в клетки. В связи с увеличенным синтезом белков в крови падает содержание аминокислот. Происходит задержка в организме азота (баланс азота становится положительным), а также фосфора, кальция, натрия.
Для эффекта соматотропина, усиливающего синтез белка в клетках, необходимо наличие углеводов и инсулина. После удаления поджелудочной железы у животных, а также при исключении из пищи углеводов действие гормона роста тормозится. Введение больших количеств этого гормона усиливает секрецию инсулина у молодых животных, у взрослых животных секреция инсулина не усиливается, а островки поджелудочной железы перерождаются, и возникает сахарный диабет. При введении гормона роста усиливаются мобилизация жира из депо и использование его в энергетическом обмене. Это ведет, к увеличению расхода жиров, также к повышению уровня кетоновых тел в крови и выделению их с мочой.
Соматотропный гормон выделяется непрерывно на протяжении всей жизни организма. Его выделение стимулируется сомтaтотропин высвобождающим фактором и тормозится соматостатином - продуктами нейросекреции гипоталамуса. У детей раннего возраста изменения, возникающие при недостаточной выработке гормона роста, проявляются в резкой задержке роста. При этом на всю жизнь человек остается. карликом (гипофизарный нанизм). Телосложение у таких людей относительно пропорционально, однако кисти и стопы малы, пальцы тонкие, окостенение скелета запаздывает, половые органы недоразвиты, вторичные половые признаки слаборазвиты, волосы отличаются мягкостью и шелковистостью, свойственной детям. Такие люди плохо переносят инфекционные и другие-болезни, часто умирают молодыми. У мужчин, страдающих этим заболеванием, отмечается импотенция, т. е. неспособность к половому акту. У женщин - стерильность, т. е. неспособность к зачатию.
При избыточной продукции гормона роста в детском возрасте развивается гигантизм; рост человека может достигать 240-250 см, а масса тела - 150 кг и более. Если же избыточная продукция гормона роста возникает у взрослого, то рост тела в целом не увеличивается, так как он уже завершен, но увеличиваются размеры тех частей тела, которые еще сохраняют способность расти: пальцев рук и ног, кистей и стоп, носа, нижней челюсти, языка, органов грудной и брюшной полостей. Это заболевание называется акромегалией. Как у гипофизарных гигантов, так и у больных акромегалией наблюдается нарушенная функция желез внутренней секреции, регулируемых гормонами передней доли гипофиза, в частности недостаточность внутрисекреторной функции половых желез.
При акромегалии отмечается также недостаточность инсулярной ткани поджелудочной железы, приводящая к сахарному диабету. Причиной акромегалии обычно является опухоль передней доли гипофиза, состоящая из ацидофильных клеток.( Г. Косицкий 1985).
Пролактин продуцируемый ацидофильными клетками передней доли гипофиза пролактин, или лютеотропный гормон, усиливает выработку молока молочными железами, а также стимулирует развитие желтого тела. Он разрушается ферментами пищеварительного тракта, поэтому его необходимо вводить в организм подкожно или внутривенно.
Если у кормящих крыс удалить гипофиз, то лактация, т. е. выделение·молока, прекращается. Введение пролактина не только усиливает отделение молока у кормящих самок, но вызывает также небольшое его отделение у некормящих, если они достигли половой зрелости и даже в том случае, если они кастрированы. Инъекции пролактина могут вызвать лактацию и у самцов. Однако для этого нужно предварительно вводить им в течение некоторого времени эстрогены и прогестерон, так как молочные железы у самцов находятся в рудиментарном состоянии и не могут лактировать, если не стимулировать искусственным путем развитие их железистой ткани. Введение пролактина даже до достижения половой зрелости вызывает формирование материнского инстинкта.
Пролактин уменьшает потребление глюкозы тканями, что вызывает повышение ее количества в крови, т. е. действует в этом отношении подобно соматотропину, однако значительно слабее. Стимуляция секреции пролактина осуществляется рефлекторно центрами гипоталамической области. Рефлекс возникает при раздражении рецепторов сосков молочных желез (во время сосания). Это приводит к возбуждению ядер гипоталамуса, которые влияют на функцию гипофиза гуморальным путем. Однако в отличие от регуляции секреции ФСГ и ЛГ гипоталамус не стимулирует, а тормозит секрецию пролактина, выделяя пролактинтормозящий фактор. Рефлекторная стимуляция секреции пролактина осуществляется путем уменьшения выработки пролактинтормозящего фактора.
Адренокортикотропный гормон (адренокортикотропин) (АКТГ) разных видов животных имеют различную структуру и отличаются по своей активности. АКТГ вызывает разрастание пучковой и сетчатой зон коры надпочечников и усиливает синтез их гормонов. Это действие АКТГ наблюдается и в том случае, если у животного предварительно был удален гипофиз и указанные зоны коры надпочечников подверглись атрофии вследствие отсутствия в организме собственного АКТГ. Удаление гипофиза не ведет к атрофии клубочковой зоны коры и мозгового вещества надпочечников.
Это говорит о том, что действие АКТГ специфично и распространяется только на пучковую и сетчатую зоны коры надпочечников.
Секреция АКТГ гипофизом усиливается при воздействии всех чрезвычайных раздражителей, вызывающих в организме состояние напряжения (стресс). Такие раздражители рефлекторно, а также вследствие повышенного выделения адреналина мозговым слоем надпочечников действуют на ядра гипоталамуса, в которых усиливается образование кортикотропинвысвобождающего фактора. Это вещество вследствие сосудистой связи гипоталамуса и гипофиза достигает клеток передней доли и стимулирует секрецию АКТГ. Последний, действуя на надпочечник, вызывает усиление выработки глюкокортикоидов (способствующих повышению сопротивляемости организма неблагоприятным факторам), а также в некоторой мере и минералокортикоидов. ( Н. Федюкович 2001).
Тиреотропный гормон (тиротропин). Выделяемый базофильными клетками передней доли гипофиза тиротропный гормон (ТТГ) стимулирует функцию щитовидной железы. Механизмы этой стимуляции многообразны. Активируя протеазы, ТТГ повышает распад тироглобулина в щитовидной железе, что приводит к усиленному выделению тироксина и трийодтиронина в кровь. ТТГ способствует накоплению йода в щитовидной железе; кроме того, он повышает активность ее секреторных клеток и увеличивает их число.
Введение ТТГ вызывает разрастание щитовидной железы, а удаление гипофиза ведет у молодых животных к ее недоразвитию, у взрослых же - к ее уменьшению и частичной атрофии. У животных после удаления гипофиза понижается основной и белковый обмен. Он может быть снова повышен введением тироксина, пересадкой гипофиза или введением тиротропина. Введение тироксина нормализует основной и белковый обмен: таким путем возмещается недостаточная продукция тироксина в атрофированной щитовидной железе животного, а пересадка гипофиза или введение тиротропного гормона нормализуют обмен, вызывая разрастание щитовидной железы, подвергшейся атрофии в отсутствие этого гормона.
Если в течение продолжительного времени вводить животным ежедневно достаточно большие количества тиротропного гормона, у них появляются симптомы, напоминающие базедову болезнь человека.
Тиротропин выделяется в небольших количествах непрерывно. Стимуляция секреции тиротропина осуществляется гипоталамусом, нервные клетки которого продуцируют тиротропинвысвобождаюший фактор, стимулирующий образование тиротропина в аденогипофизе. Уровень секреции тиротропина зависит от количества гормонов щитовидной железы в крови. При достаточном количестве последних секреция тиротропина угнетается. Недостаточное содержание в крови гормонов щитовидной железы, наоборот, стимулирует секрецию тиротропина. Таким образом, и здесь функционирует механизм обратной связи.
При охлаждении организма секреция тиротропина усиливается и увеличивается образование гормонов щитовидной железы, в результате чего повышается продукция тепла.
Если организм подвергается повторному действию охлаждения, то стимуляция секреции тиротропина возникает даже при действии сигналов, предшествующих охлаждению, вследствие возникновения условных рефлексов. Отсюда следует, что кора головного мозга может
8-09-2015, 23:52