СОДЕРЖАНИЕ
1. Введение ……………………………………………….. 3 стр.
2. История распространения холеры …………………… 3 стр.
3. Морфология. Типичные вибрионы Эль-Тор ………… 5 стр.
3.1. Морфология ……………………………………….. 5 стр.
3.2. Клеточные структуры холерного вибриона …….. 6 стр.
3.3. Тинкториальные свойства ……………………….. 7 стр.
3.4. Культуральные свойства ………………………… 7 стр.
4. Рост возбудителя холеры ………………………………. 8 стр.
5. Биохимические свойства холеры. Биовары холеры ….. 9 стр.
6. Патогенез ……………………………………………….. 11 стр.
7. Токсины холерных вибрионов ………………………... 11 стр.
8. Клинические проявления холеры ……………………… 12 стр.
9. Лабораторная диагностика …………………………….. 12 стр.
9.1 Ускоренные методы диагностики ……………….. 13 стр.
9.2 Дифференциальная диагностика ………………… 13 стр.
10. Лечение ………………………………………………... 13 стр.
11. Иммунитет …………………………………………….. 14 стр.
12. Список литературы …………………………………… 15 стр.
1. ВВЕДЕНИЕ.
Холера - острая кишечная инфекция, вызывается Vibrio cholerae и характеризуется развитием гастроэнтерита, сопровождающегося обезвоживанием организма и нарушением электролитного кислотно-щелочного баланса, склонна к эпидемическому и пандемическому распространению. По определению ВОЗ, холера - это болезнь, для которой типичен острый тяжелый обезвоживающий понос с испражнениями в виде рисового отвара.
В связи со способностью в короткие сроки поражать обширные контингенты населения холера относится к болезням, на которые распространяются Международные медико-санитарные правила как на конвенционные, особо опасные и карантинные инфекции.
Выделяют два типа эпидемий холеры:
1 тип: эпидемии-вспышки с едиными источником инфекции и путями распространения, характеризующиеся одномоментным появлением большого количества больных,
2 тип: вялотекущие эпидемии с перманентной заболеваемостью небольшого контингента и трудно выявляемыми путями передачи возбудителя.
2. ИСТОРИЯ РАСПРОСТРАНЕНИЯ ХОЛЕРЫ.
Еще до нашей эры были описаны характерные признаки данного заболевания, указывались на быстрое распространение и высокую летальность. У истории холеры принято условно выделять несколько периодов:
Первый период охватывает промежуток от древнейших времен до 1817 г, в течении которого заболевание было эндемичным для районов Юго-Восточной Азии, особенно Бенгалии т.е. дельты Ганга и Брахмапутры, считающейся "колыбелью холеры".
Второй период исчисляется с 1817 по 1926 гг., когда в связи с расширением международных экономических связей и колониальными войнами (в первую очередь колониальной экспансией Великобритании в Индии и на Среднем Востоке) стало возможным пандемическое распространение холеры. В этот период (1823-1926) Россия перенесла 57 холерных лет. С 1817 года по 1926 год на земном шаре переболело холерой 4.5 млн. человек, из них 2 млн. скончались от холеры. В России за это время значительные вспышки холеры случались 8 раз. От холеры умер П.И.Чайковский.
В Западную Европу и Россию холера проникла во время польского восстания 1830-1831 гг. Основными воротами для прорыва возбудителя в Европу были некоторые регионы Ближнего Востока, Египет и порты Средиземноморья. Первым описал возбудителя заболевания итальянец Пачини и независимо от него врач Недзвецкий.
В чистой культуре микроб выделен в ходе экспедиций в Египет (1883-1884) Робертом Кохом ("запятая Коха") подробно описавшем его свойства. В экспедициях участвовали французские (Ру, Нокар, Штраусс, Тюйе) и немецкие (Кох, Гаффки, Фишер, Тресков) ученые. Работа исследователей проходила в сложной и нервной обстановке, подогреваемой кампанией франко-прусского шовинизма. Во время исследований Луи Тюйе заразился и погиб. Роберт Кох возложил на могилу ученого венок со словами: "Это скромный венок из лавров, но им венчают героев".
В третий период (1926-1961) возбудитель локализовался в Юго-Восточной Азии (Пакистан, Индия, Бангладешь). В 1939 г на острове Суловеси (Индонезия) и ряде других стран Юго-Восточной Азии были описаны эпидемические вспышки холеры, вызванные особым биологическим вариантом холерного вибриона Эль-Тор, который в дальнейшем получил всемирное распространение в ходе VII пандемии холеры. Вибрион Эль-Тор был выделен еще в 1906 году на карантинной станции Эль-Тор в Египте, супругами Готшлих из трупов мусульман-поломников, но поскольку в то время эпидемии не было, то роль вибрионов Эль-Тор осталась недоказанной.
В четвертый исторический период холеры, начавшийся в 1961 г был обусловлен вибрионом Эль-Тор и О139. Серогруппа О139 была выделена в 1992 году на юге Индии, тогда заболевание расценивалось как холероподобное, т.к. было вызвано пока еще неизвестным вибрионом.
В СССР холера проникла из Афганистана в 1965 году, наблюдались вспышки холеры в Каракалпакии, в Узбекистане. Россия вступила в VII пандемию холеры в 1970 году, когда заболевание вспыхнуло в Астраханской области, Заволжье и Одессе. Тогда информация о приближающейся инфекции стала известна от моряков, санитарные службы быстро отреагировали на известие, были своевременно созданы госпиталя для больных, порты и дороги закрыты на карантин.
В начале 1993 г. появились сообщения о вспышках холеры в юго-восточной Азии, вызванных вибрионами ранее неизвестной серогруппы, обозначенных как серовар холеры 0139 (Бенгал). Произошла смена возбудителя с классического вибриона на вибрион Эль-Тор. При этом болезнь стала протекать относительно доброкачественно. Доля смертельных исходов уменьшилась.
В связи с широким распространением холеры и постоянной угрозой завоза на территорию Российской Федерации, проблема этого заболевания продолжает сохранять свою актуальность . Состояние заболеваемости холерой в мире в настоящее время определяется эпидемиями и вспышками инфекции в Африке, где в 23 странах сформировались эндемичные очаги холеры. В 2003 г. во Всемирную организацию здравоохранения сообщили 29 стран о 91865 больных холерой, из которых 1485 человек умерли. Крупные вспышки холеры в 2003 году были отмечены в Либерии (33604 больных), Конго (22768 больных), Мозамбике (13758 больных), Сомали (4877 больных), Уганде (3433 больных), Индии (2025 больных).
Еще одна особенность последней пандемии - часто остается вибриононосительство. Регистрируется на всех континентах. Максимальная заболеваемость в последние годы в странах Центральной и Южной Америки. Вспышки холеры растянулись во времени - период повышенной заболеваемости стал более длительным и медленно идет на спад, нет пиков заболеваемости.
3. МОРФОЛОГИЯ. ТИПИЧНЫЕ ВИБРИОНЫ ЭЛЬ-ТОР.
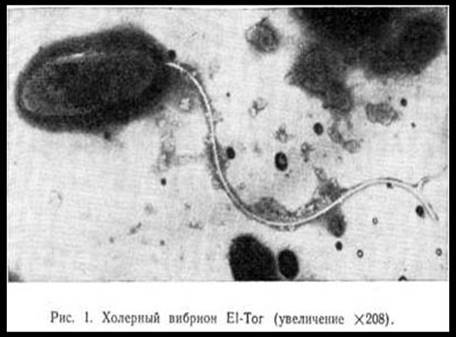
3.1. МОРФОЛОГИЯ. Типичные вибрионы Эль-Тор, так же как и вибрионы азиатской холеры, представляют собой короткие изогнутые палочки от 1,5 до 3 мк в длину и от 0,2 до 0,6 мк в ширину (рис. 1 И 2). Вибрионы отличаются значительным полиморфизмом: наряду с типичными формами в препаратах из содержимого кишечника реконвалесцентов и лабораторных (особенно старых бульонных) культур могут встречаться малоизогнутые палочкообразные, S-образные, спиралевидные, нитевидные, кокковидные, дрожжеподобные и гигантские формы. Такие нетипичные формы холерных вибрионов, |

как правило, образуются при их выращивании в неблагоприятных условиях: на средах, содержащих литий, крахмал, гликокол, иммунную сыворотку, избыточную концентрацию поваренной соли и т. Д. Спор и капсул холерные вибрионы не образуют, поэтому их устойчивость во внешней среде сравнительно не велика.
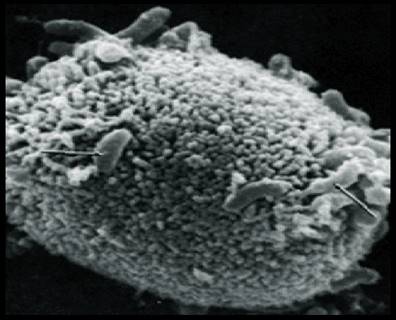
Как показали исследования Фоллет и Гордон (1963), Куно и Огасавара (1964), Галлю (1969), а также отечественных авторов (Л. Ф. Зыкин, 1967; Л. Ф. Зыкин и Л. С. Петрова, 1967), клетка холерного вибриона аналогична по строению с грамотрицательными бактериями: имеет клеточную стенку или оболочку, цитоплазматическую Мембрану, цитоплазму с включениями, жгутик и ворсинки, отходящие от наружного слоя оболочки. Последняя представлена тремя слоями, пронизанными канальцами, через которые совершаются процессы обмена, в частности выделяются токсические субстанции, вырабатываемые клеткой. По данным Миера и Шривастава (1959, 1961), соматический антиген находится в клеточной оболочке и цитоплазматической мембране, которая участвует в процессах движения, а также в синтезе белка и полирибонуклеотидов.
Холерные вибрионы относятся к группе активно подвижных бактерий. Более того, они, по-видимому, являются самыми отличными «бегунами» из всех групп подвижных микробов. Их подвижность обусловлена наличием мощного и длинного жгутика на одном из концов клетки. Следует отметить, что определение подвижности холерных вибрионов является важным диагностическим признаком, поэтому изучение так называемой висячей или раздавленной капли — один из обязательных тестов при бактериологической диагностике холеры. Исключительная подвижность холерного вибриона, как показали исследования Куно и Огасавара, обусловлена наличием особой мембраны, идущей вдоль жгутика, которая напоминает ундулирующую мембрану простейших.
3.2. КЛЕТОЧНЫЕ СТРУКТУРЫ ХОЛЕРНОГО ВИБРИОНА.
Клеточные структуры холерного вибриона не равнозначны в антигенном отношении. Наиболее иммуногенными являются оболочка и жгутик вибриона. Характерной чертой антигенной структуры вибрионов азиатской холеры, отличающей их от Эль-Тор, как показали исследования Тамаоки с сотрудниками (1967) и Щурки- ной (1969), является содержание в клетках Эль-Тор фактора S3, а в вибрионах азиатской холеры — Sb. При подтверждении результатов этих исследований откроются дополнительные возможности по серологическому типированию холерных вибрионов.
3.3. ТИНКТОРИАЛЬНЫЕ СВОЙСТВА.
Холерные вибрионы интенсивно окрашиваются всеми основными анилиновыми красителями. Наиболее широко распространена окраска препаратов разведенным (1:10) карболовым фуксином Циля, при которой вибрионы приобретают интенсивно- розовый цвет.
Холерные вибрионы, как, впрочем, и все близкие им по первичной локализации в инфицированном организме, а отсюда и по механизму передачи инфекции патогенные представители семейства кишечных относятся к большой группе грамотрицательных микроорганизмов. Иными словами, они не воспринимают окраску по Граму в связи с отсутствием в соме их клеток магниевых солей рибонуклеиновой кислоты.
Для окраски жгутика холерных вибрионов используются специальные методы, в большинстве случаев включающие «протраву» органоидов движения солями тяжелых металлов (серебрение). 
3.4. КУЛЬТУРАЛЬНЫЕ СВОЙСТВА.
Холерный вибрион предпочитают аэробные условия и быстро погибает в анаэробных. Температурный оптимум 37 °С. Вибрион холеры хорошо растёт на простых питательных средах с высоким рН (7,6-8,0); подобные галофильные свойства используют в подборе элективных средств.
На твёрдых средах возбудитель холеры образует небольшие круглые дисковидные прозрачные S-колонии с ровными краями, голубоватые в проходящем свете, что сразу их отличает от более грубых и мутно-белых колоний энтеробактерий.
Старые культуры холеры несколько грубеют и приобретают желтовато-коричневый оттенок. На агаре с тиосульфатом, цитратом, солями жёлчных кислот и сахарозой (TCBS-arap) V. cholerae ферментирует последнюю и образует жёлтые колонии. При посеве уколом в желатину через 48-72 ч микроорганизм дает воронкообразное разжижение, верхняя часть которого при просматривании сбоку представляется пузырьком воздуха. Позднее разжижение увеличивается, полость заполняется белёсой массой вибрионов.
Холерный вибрион также образует неровные мутные R-колонии; бактерии, из которых не чувствительны к бактериофагам, антибиотикам и не агглютинируются О-антисыворотками.
В жидких средах вибрион холеры вызывает помутнение и образование нежной голубоватой плёнки на поверхности, её края приподняты вдоль стенок пробирки; при встряхивании она легко разрушается и оседает на дно.
4. РОСТ ВОЗБУДИТЕЛЯ ХОЛЕРЫ.
Холерные вибрионы относятся к так называемой группе галофилов, иными словами, щелочелюбивых микроорганизмов. Оптимальный показатель концентрации водородных ионов в среде выращивания 7,6—8,0, однако хороший рост наблюдается и при рН 9,2. Это свойство холерных вибрионов широко используется при подборе элективных питательных сред для бактериологической диагностики: в высокощелочной среде (например, в щелочной пептонной воде) уже к 4—6-му часу после посева кала больного холерой регистрируется обильный рост вибрионов, тогда как множество других представителей семейства кишечных, как патогенных, так и комменсалов, размножается при этом крайне скудно. С другой стороны, галофильность холерных вибрионов определяет их очень высокую чувствительность к действию кислот, что может быть использовано при дезинфекции различных объектов в очагах холеры и, что особенно важно, при обеззараживании питьевой воды.
На 1 щелочной пептонной воде при рН 7,8 уже через 6—8 часов после посева холерные вибрионы образуют нежную и не всегда хорошо заметную голубовато- серую пленку, края которой приподняты вдоль стенок пробирки; остальная часть среды лишь слегка диффузно мутнеет. При встряхивании пленка легко разрушается и оседает на дно пробирки. Пептонная вода с добавлением 0,5-1% NaCl — наилучшая среда накопления для возбудителя холеры.
5. БИОХИМИЧЕСКИЕ СВОЙСТВА ХОЛЕРЫ. БИОВАРЫ ХОЛЕРЫ.
Вибрион холеры классический (биотип холера), и вибрион холеры биотип Эль-Тор отличаются по биохимическим свойствам.
Холерные вибрионы обладают широким спектром биохимической активности: наличие набора протеолитических ферментов обусловливает их способность разжижать желатину и образовывать индол на средах с пептоном. Холерные вибрионы разлагают до кислоты (без газа) сахарозу, глюкозу, мальтозу, маннозу, маннит, лактозу и благодаря наличию фермента диастазы (Brieger, Fermi) расщепляют крахмал, восстанавливают нитраты в нитриты.
Как подчеркивалось ранее, варианту (биотипу) Эль-Тор присуща способность гемолизировать эритроциты барана (козы), тогда как классические варианты, как правило, не продуцируют гемолитический фермент. Биохимические свойства холерных вибрионов используют как для их дифференциации от большого числа не патогенных для человека водных вибрионов, так и для внутривидового типирования. Так, определение сахаролитических ферментов было использовано Хейбергом (1934, 1935) для подразделения всех вибрионов на 6 групп, совпадающих с группами установленными Гарднером и Венкатраменом на основании изучения антигенной структуры всех микроорганизмов. 
По данным Хейберга, патогенные для человека классические и Эль-Тор вибрионы, обладающие в отличие от непатогенных вибрионов I типом ферментации (постоянно разлагают маннозу и сахарозу и инертны в отношении арабинозы), относятся к I—О серологической группе Гарднера и Венкатрамена, как известно, включающей в свой состав возбудителей холеры. Способность холерных вибрионов восстанавливать нитраты в нитриты и продуцировать индол используется при постановке так называемой Cholerarot реакции. Эта реакция, предложенная Пелем и Буйдвидом, очень постоянная, хотя и не строго специфичная для холерных вибрионов, регистрируется в виде розовато-красного окрашивания культур, выращенных на пептонной воде после добавления нескольких капель серной или соляной кислоты. Механизм этой реакции следующий: добавленная кислота вытесняет из восстановленных холерными вибрионами нитритов азотистую кислоту, которая соединяется с индолом и образует нитрозоиндол — вещество ярко-красного цвета.
Гемолитические свойства вибрионов Эль-Тор, впервые описанные Koch и Bitter, используются для их дифференциации от классического биотипа возбудителей холеры. Холерные вибрионы сбраживают с образованием кислоты многие углеводы (глюкозу, сахарозу, мальтозу, маннит, лактозу, гликоген, крахмал и др.). Ферментация маннозы, сахарозы и арабинозы (так называемая триада Хёйберга) имеет диагностическое значение.
По способности разлагать эти три углевода все вибрионы холеры разделяют на 6 групп. Холерные вибрионы разлагают только маннозу и сахарозу и принадлежат к 1-й группе Хейберга.
Бактерии холеры этой группы обладают плазмокоагулирующим (свёртывают плазму кролика) и фибринолитическим (разжижают свёрнутую сыворотку по Леффлеру) свойствами.
Холерные вибрионы свёртывают молоко и разлагают другие белки и их дериваты до аммиака и индола; H2S не образуют, восстанавливают нитраты и образуют индол (эту способность учитывают в нитрозоиндоловой реакции, также известной как холера-рот реакция).
На основании биохимических и биологических различий холерные вибрионы разделяют на два биовара — классический (V. cholerae биовар asiaticae) и Эль-Тор (V. cholerae биовар eltor).
Бактерии серовара О139 (Бенгал) устойчивы к полимиксину и не проявляют гемолитической активности.
Дифференциальные признаки возбудителей холеры
| Признак | V. cholerae биовар asiaticae | V. cholerae биовар eltor | серовара О139 (Бенгал) |
| Реакция Фогеса-Проскауэра | + (чаще - ) | + (чаще + ) | + (чаще + ) |
| Чувствительность к полимиксину В (50ЕД) | + | - | - |
| Гемолиз эритроцитов барана | - | + | - |
| Агглютинация куриных эритроцитов | - | + (чаще +) | + (чаще + ) |
| Чувствительность к классическому монофагу | + | - | - |
| Чувствительность к монофагу Эль-Тор | - | + | - |
| Гексаминовый тест | - | + | - |
6. ПАТОГЕНЕЗ.
Проникнув через рот часть вибрионов в желудке погибает. Если есть предрасполагающие факторы, повышенное потребление жидкости, то вибрионы проникают в кишечник. Там щелочная среда и высокое содержание пептонов (много молекул белки и других питательных веществ). Это способствует интенсивному размножению вибрионов. Выделение экзотоксина и факторов проницаемости приводит к основным клиническим проявлениям.
Холероген является пусковым механизмом патологического процесса. Действию холерогена помогает фактор проницаемости - ферменты типа нейраминидазы. Нейраминидаза расщепляет гликопротеиды. В мембране энтероцитов содержатся ганглиозиды G и М. Видоизменение этого ганглиозида и само присутствие холерогена приводит к тому, что активируется фермент аденилатциклаза. Это приводит к усилению синтеза цАМФ (концентрация в энтероците возрастает в десятки, сотни раз) цАМФ - это регуляторное вещество которое меняет потоки жидкости. В норме функцией энтероцита является всасывание жидкости из просвета кишечника внутрь клетки. Высокая концентрация цАМФ приводит к тому, что жидкость из клетки секретируется в просвет кишечника. Изучение сока кишечника в этом случае показывает, что в 1 литре содержится 5 г поваренной соли, 4 г. гидрокарбоната натрия и 1 г. калия хлорида. Это соотношение очень важно для лечения. Соотношение электролитов в 1 л кишечного сока 5 к 4 к 1 можно вводиться внутривенно капельно, а можно давать перорально вместе с сахарами. Сахара способствуют усвоению электролитов. Поскольку жидкости теряется очень много то развивается гиповолемия (уменьшение ОЦК). Повышается вязкость крови. Развивается сосудистая недостаточность. Недостаток кислорода в тканях. Из-за недостатка кислорода происходит нарушение метаболизма, развивается ацидоз. В последующем грубые метаболические нарушения могу приводить к некрозу почечных канальцев. С водой уходят электролиты. Наиболее существенна потеря калия. Гипокалиемия приводит к развитию мышечной слабости, а отдельные мышечные группы могут некоординированно сокращаться что ведет к появлению судорожного синдрома.
7. ТОКСИНЫ ХОЛЕРНЫХ ВИБРИОНОВ.
В ответ на проникновение бактерий холеры эпителиальные клетки выделяют щелочной секрет, служащий идеальной средой для размножения возбудителя. Основной фактор патогенности — способность к токсинообразованию. Холерные вибрионы образуют эндо-и экзотоксины.
Эндотоксин холеры — термостабильный ЛПС, сходный по структуре и активности с эндотоксинами прочих грамотрицательных бактерий. Он проявляет иммуногенные свойства, индуцируя синтез вибриоцидных AT.
Однако этот токсин холеры не играет существенной роли в развитии характерных проявлений.
Экзотоксин холеры
(холероген) — термолабильный белок; его образование кодируют как хромосомные, так и
8-09-2015, 23:56