2. Гены в хромосоме расположены линейно, каждый ген занимает в хромосоме определенный локус (место).
3. Гены , расположенные в одной хромосоме, образуют группу сцепления и вместе (сцеплено) передаются потомкам, число групп сцепления равно гаплоидному набору хромосом.
4. Сцепление не абсолютно, так как в профазе мейоза может происходить кроссинговер и гены, находящиесяв одной хромосоме, разобщаются. Сила сцепления зависит от расстояния между генами в хромосоме: чем больше расстояние, тем меньше сила сцепления. и наоборот. Расстояние между генами измеряется в процентах кроссинговера. 1% кроссинговера соответтствует одной морганиде.[4]
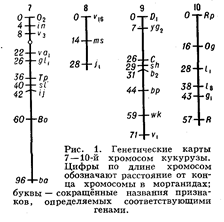
Генетические карты хромосом составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, по мере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые назв. мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры . Составить Генетические карты хромосом можно только для объектов, у которых изучено большое число мутантных генов. Например, у дрозофилы идентифицировано свыше 500 генов, локализованных в её 4 группах сцепления, у кукурузы — около 400 генов, распределённых в 10 группах сцепления (рис. 1). У менее изученных объектов число обнаруженных групп сцепления
меньше гаплоидного числа хромосом. Так, у домовой мыши выявлено около 200 генов, образующих 15 групп сцепления (на самом деле их 20); у кур изучено пока всего 8 из 39. У человека из ожидаемых 23 групп сцепления (23 пары хромосом) идентифицировано только 10, причём в каждой группе известно небольшое число генов; наиболее подробные карты составлены для половых хромосом.
|
|
|
|

|
|
У бактерий, к-рые являются гаплоидными организмами, имеется одна, чаще всего непрерывная, кольцевая хромосома и все гены образуют одну группу сцепления (рис. 2). При переносе генетич. материала из клетки-донора в клетку-реципиент, например при конъюгации , кольцевая хромосома разрывается и образующаяся линейная структура переносится из одной бактериальной клетки в другую (у кишечной палочки в течение 110-120 мин). Искусственно прерывая процесс конъюгации, можно по возникшим типам рекомбинантов установить, какие гены успели перейти в клетку-реципиент. В этом состоит один из методов построения Генетических карт хромосом бактерий, детально разработанных у ряда видов. Ещё более детализированы Генетические карты хромосом нек-рых бактериофагов
Генетика пола . Количество групп сцепленных генов оказалось равным количеству пар хромосом, присущих данному виду. Важнейшие доказательства хромосомной теории наследственности были получены при изучении наследования, сцепленного с полом . Ранее цитологи открыли в хромосомных наборах ряда видов жиивотных особые , так называемые половые хромосомы , которыми самки отличаются от самцов. В одних случаях самки имеют 2 одинаковые половые хромосомы(XX), а самцы-разные(XY), в других - самцы-2 одинаковые(XX, или ZZ), а самки - разные(XY, или ZW). Пол с одинаковыми половыми хромосомами называется гомогаметным , с разными - гетерогаметным . Женский пол гомогаметен , а мужской гетерогаметен у некоторых насекомых ( в том числе у дрозофилы) и всех млекопитающих. Обратное соотношение - у птиц и бабочек. Ряд признаков у дрозофилы наследуется в
строгом соответствии с передачей потомству X-хромосом. Самка дрозофилы, проявляюща
рецесивный признак , например белую окраску глаз, в силу гомозиготности по этому гену, находящимуся в X-хромосоме, передает белую окраску глаз всем сыновьям, так как они получают свою X-хромосому только от матери. В случае гетерозиготности по рецессивному сцепленному с полом признаку самка передает его половине сыновей. При противоположном определении пола (самцы XX, или ZZ; самки-XY, или ZW) особи мужского пола передают сцепленные с полом признаки дочерям, получающим свою X( =Z ) хромосому от отца. Иногда в результате нерасхождения половых хромосом при мейозе возникают самки строения XXY и самцы XYY. Возможны также случаи соединения X-хромосом концами; тогда самки передают сцепленные X-хромосомы своим дочерям, у которых и проявляются сцепленные с полом признаки. Сыновья же похожи на отцов (такое наследование называется гологеническим ). Если наследуемые гены находятся в Y-хромосоме, то определяемые ими признаки передаются только по мужской линии - от отца к сыну (такое наследование называется голандрическим ). Хромосомная теория наследственности вскрыла внутриклеточные механизмы наследственности, дала точное и единое объяснение всех явлений наследования при половом размножении, объяснила сущность изменений наследственности, то есть изменчивости.
Нехромосомная теория наследственности . Первенствующая роль ядра и хромосом в наследственности не исключает передачи некоторых признаков и через цитоплазму, в которой обнаружены структуры, способные к самовоспроизведению.Единицы цитоплазматической (нехромосомной) наследственности отличаются от хромосомных тем, что они не расходятся при мейозе. Поэтому потомство при нехромосомной наследственности воспроизводит признаки только одного из родителей (чаще матери ). Таким образом , различают ядерную наследственность , связанную с передачей наследственных признаков, находящихся в хромосомах ядра (иногда ее называют хромосомной наследственностью ), и внеядерную наследственность , зависящую от передачи самовоспроизводящихся структур цитоплазмы. Ядерная наследственность реализуется и при вегетативном размножении , но не сопровождается перераспределением генов, что наблюдается при половом размножении, а обеспечивает константную передачу признаков из поколения в поколение, нарушаемую только соматическими мутациями .
Молекулярная генетика . Применение новых физических и химических методов, а также использование в качестве объектов исследования бактерий и вирусов резко повысили разрешающую способность генетических экспериментов, привели к изучению наследственности на молекулярном уровне и бурному развитию молекулярной генетики . Впервые Н. К. Кольцов (1927 г) выдвинул и обосновал представления о молекулярной основе наследственности и о матричном способе размножения “наследственных молекул”.В 40-х гг. 20 в. была экспериментально доказана генетическая роль дизоксирибонуклеиновой кислотиы ( ДНК ) , а в 50-60-х гг. установлена ее молекулярная структура и выяснены принципы кодирования генетической информации. Генетическая информация ,заложенная в наследственных структурах организмов (в хромосомах, цитоплазме, клеточных организмах), получаемая от предков в виде совокупности генов информация о составе, строении и характере обмена составляющих организм веществ (прежде всего белков и нуклеиновых кислот) и связанных с ними функциях. У многоклеточных форм при половом размножении генетическая информация передаётся из поколения в поколение через посредство половых клеток — гамет , единственная функция к-рых — передача и хранение генетической информации. У микроорганизмов и вирусов имеются особые типы ее передачи . Генетическая информация заключена преимущественно в хромосомах, где она зашифрована в определённой линейной последовательности нуклеотидов в молекулах дезоксирибонуклеиновой кислоты — ДНК (генетический код). Генетический код - это система зашифровки наследственной информации в молекулах нуклеиновых кислот, реализующаяся у животных, растений, бактерий и вирусов в виде последовательности нуклеотидов . В природных нуклеиновых кислотах — дезоксирибонуклеиновой (ДНК) и рибонуклеиновой (РНК)—встречаются 5 распространённых типов нуклеотидов (по 4 в каждой нуклеиновой к-те), разлчающихся по входящему в их состав азотистому основанию . В ДНК встречаются основания:
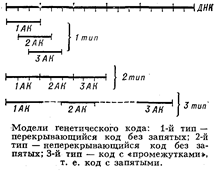
аденин (А), гуанин (Г), цитозин (Ц), тимин (Т); в РНК вместо тимина присутствует урацил (У). Кроме них, в составе нуклеиновых к-т обнаружено ок. 20 редко встречающихся (т. н. неканонических, или минорных) оснований, а также необычных Сахаров. Так как количество кодирующих знаков Генетического кода (4) и число разновидностей аминокислот в белке (20) не совпадают, кодовое число (т. е. кол-во нуклеотидов, кодирующих 1 аминокислоту) не может быть равно 1. Различных сочетаний по 2 нуклеотида возможно лишь 42 = 16, но этого также недостаточно для зашифровки всех аминокислот. Американский учёный Г. Гамов предложил (1954) модель т р и п л е т н о г о генетического кода, т. е. такого, в котором 1 аминокислоту кодирует группа из трёх нуклеотидов, называемых кодоном. Число возможных триплетов равно 43 = 64, а это более чем втрое превышает число распространённых аминокислот, в связи с чем было высказано предположение, что каждой аминокислоте соответствует несколько кодонов (так называемая вырожденность кода). Было предложено много различных моделей генетического кода, из которых серьёзного внимания заслуживали три модели (см. рис.): перекрывающийся код без запятых, неперекрывающийся код без запятых и код с запятыми. В 1961 Ф. Крик (Великобритания) с сотрудниками получил подтверждение гипотезы триплетного неперекрывающегося кода без запятых. Установлены след. осн. закономерности, касающиеся генетического кода: 1) между последовательностью нуклеотидов и кодируемой последовательностью аминокислот существует линейное соответствие (коллинеарность генетического кода); 2) считывание кода начинается с определённой точки; 3) считывание идёт в одном направлении в пределах одного гена; 4) код является неперекрывающимся; 5) при считывании не бывает промежутков (код без запятых); 6) генетический код, как правило, является вырожденным, т. е. 1 аминокислоту кодируют 2 и более триплетов-синонимов (вырожденность генетического кода уменьшает вероятность того, что мутационная замена основания в триплете приведёт к ошибке); 7) кодовое число равно трём;
|
|
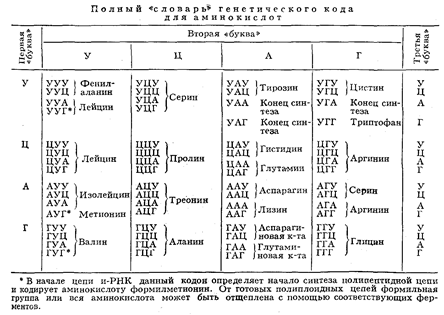
8) код в живой природе универсален (за нек-рыми исключениями). Универсальность генетического кода подтверждается экспериментами по синтезу белка in vitго. Если в бесклеточную систему, полученную из одного организма (например, кишечной палочки), добавить нуклеиновокислотную матрицу, полученную из другого организма, далеко отстоящего от первого в эволюционном отношении (например, проростков гороха), то в такой системе будет идти белковый синтез. Благодаря работам амер. генетиков М. Ниренберга, С. Очоа, X. Корана известен не только состав, но и порядок нуклеотидов во всех кодонах..
Из 64 кодонов у бактерий и фагов 3 кодона — УАА, УАГ и УГА — не кодируют аминокислот; они служат сигналом к освобождению полипептидной цепи с рибосомы , т. е. сигнализируют о завершении синтеза полипептида. Их наз. терминирующими кодонами. Существуют также 3 сигнала о начале синтеза — это т. н. инициирующие колоны — АУГ, ГУГ и УУГ,— к-рые, будучи включёнными в начале соответствующей информационной РНК (и-РНК), определяют включение формилметионина в первое положение синтезируемой полипептидной цепи. Приведённые данные справедливы для бактериальных систем; для высших организмов многое ещё не ясно. Так, кодон УГА у высших организмов может быть значащим; не совсем понятен также механизм инициации полипептида.
Реализация генетического кода в клетке происходит в два этапа. Первый из них протекает в ядре; он носит назв. транскрипции и заключается в синтезе молекул и-РНК на соответствующих участках ДНК. При этом последовательность нуклеотидов ДНК « переписывается » в нуклеотидную последовательность РНК. Второй этап — трансляция — протекает в цитоплазме, на рибосомах; при этом последовательность нуклеотидов и-РНК переводится в последовательность аминокислот в белке; этот этап протекает при участии транспортной РНК (т-РНК) и соответствующих ферментов.
Генетическая информация реализуется в ходе онтогенеза — развития особи — ее передачей от гена к признаку. Все клетки организма возникают в результате делений единственной ис
ходной клетки — зиготы — и потому имеют один и тот же набор генов — потенциально одну и ту же генетическую информацию. Специфичность клеток разных тканей определяется тем, что в них активны разные гены, т. е. реализуется не вся информация, а только её часть, необходимая для функционирования данной ткани.
По мере изучения наследственности на субклеточном и молекулярном уровне углублялось и уточнялось представление о гене. Если в опытах по наследованию различных признаков ген постулировался как элементарная неделимая единица наследственности, а в свете данных цитологии его рассматривали как изолированный участок хромосомы, то на молекулярном уровне ген-входящий в состав хромосомы участок молекулы ДНК , способный к самовоспроизведению и имеющий специфическую структуру, в которой закодирована программа развития одного или нескольких признаков организма. В 50-х гг. на микроорганизмах (американский генетик С. Бензер)было показано , что каждый ген состоит из ряда различных участков, которые могут мутировать и между которыми может происходить кроссинговер. Так подтвердилось представление о сложной структуре гена, развивавшееся еще в 30-х гг. А. C. Серебровским и Н. П. Дубининым на основе данных генетического анализа.
В 1967-69 гг. был осуществлен синтез вирусной ДНК вне организма, а также химический синтез гена дрожжевой аланиновой транспортной РНК. Новой областью исследования стала наследственность соматических клеток в организме и в культурах тканей. Открыта возможность экспериментальной гибридизации соматических клеток разных видов. В связи с достижениями молекулярной биологии явления наследственности приобрели ключевое значение для понимания ряда биологических процессов, а также для множества вопросов практтики.
Наследственность и эволюция . Еще Дарвину было ясно значение наследственности для эволюции организмов. Установление дискретной природы наследственности устранило
одно из важных возражений против дарвинизма: при скрещивании особей , у которых появились наследственные изменения, последние должны якобы “ разбавляться “ и ослабевать в своем направлении. Однако, в соответствии с законами Менделя,они не уничтожаются и не смешиваются, а вновь проявляются в потомстве в определенных условиях. В популяциях яв-
ления наследственности предстали как сложные процессы, основанные на скрещиваниях между особями, отборе, мутациях , генетико-автоматических процессах и др. На это впервые указал С. С. Четвериков (1926 г.) , экспериментально доказавший накопление мутаций внутри популяции. И. И. Шмальгаузен (1946 г.) выдвинул положение о “ мобилизационном ре
зерве наследственной изменчивости “ как материале для творческой деятельности естественного отбора при изменении условий внешней среды. Показано значение разных типов изменений наследственности в эволюции. Эволюция понимается как постепенное и многократное изменение наследственности вида. в то же время наследственность, обеспечивающая постоянство видовой организации, -это коренное свойство жизни, связанное с физико-химической структкрой элементарных единиц клетки, прежде всего ее хромосомного аппарата, и прошедшее длительный период эволюции.
Принципы организации этой структуры (генетический код), по-видимому, универсальны для всех живых существ и рассматриваются как важнейший атрибут жизни.
Под контролем наследственности находится и онтогенез, начинающийся с оплодотворения яйца и осуществляющийся в конкретных условиях среды. Отсюда различие между совокупностью генов, получаемых организмом от родителей, — генотипом и комплексом признаков организма на всех стадиях его развития — фенотипом . Роль генотипа и среды в формировании фенотипа может быть различна.
Но всегда следует учитывать генотипически обусловленную норму реакции организма на влияния среды. Изменения в фенотипе не отражаются адекватно на генотипич. структуре половых клеток, поэтому традиционное представление о наследовании приобретённых признаков отвергнуто, как не имеющее фак-тич. основы и неправильное теоретически. Механизм реализации наследственности в ходе развития особи, по-видимому, связан со сменой действия разных генов во времени и осуществляется при взаимодействии ядра и цитоплазмы, в к-рой происходит синтез тех или иных белков на основе программы, записанной в ДНК и передающейся в цитоплазму с информационной РНК.
Закономерности наследственности имеют огромное значение для практики сельского х-ва и медицины. На них основываются выведение новых и совершенствование существующих сортов растений и пород животных. Изучение закономерностей наследственности привело к научному обоснованию применявшихся ранее эмпирически методов селекции и к разработке новых приёмов (экспериментальный мутагенез , гетерозис , полиплоидия и др.).
Генетика человека - это отрасль генетики, тесно связанная с антропологией и медициной. Генетика человека условно подразделяют на антропогенетику, изучающую наследственность и изменчивость нормальных признаков человеческого организма, и генетику медицинскую, к-рая изучает его наследственную патологию (болезни, дефекты, уродства' и др.). Генетика человека связана также с эволюционной теорией, так как исследует конкретные механизмы эволюции человека и его место в природе, с психологией, философией, социологией. Из направлений Генетика человека интенсивно развиваются питогенетика, биохимич. генетика, иммуногенетика, генетика высшей нервной деятельности, физиологич. генетика.
В Генетике человека вместо классич. гибридологического анализа
применяют генеалогический
метод
, к-рый состоит в анализе распределения в семьях (точнее, в родословных) лиц, обладающих данным признаком (или аномалией) и не обладающих им, что раскрывает тип наследования, частоту и интенсивность проявления признака и т. д. При анализе семейных данных получают также цифры эмпирического риска
, т. е. вероятность обладания признаком в зависимости от степени родства с его носителем. Генеалогич. методом уже показано, что более 1800 морфологич., биохимич. и др. признаков человека наследуется по законам Менделя . Например, тёмная окраска кожи и волос доминирует над светлой; пониженная активность или отсутствие нек-рых ферментов определяется рецессивными генами, а рост, вес, уровень интеллекта и ряд др. признаков — «полимерными» генами, т. е. системами из мн. генов. Мн. признаки и болезни человека, наследующиеся сцепленно с полом, обусловлены генами, локализованными в Х- или У- хромосоме. Таких генов известно ок. 120. К ним относятся гены гемофилии А и В, недостаточности фермента глюко-зо-6-фосфат-дегидрогеназы, цветовой слепоты и др. Др. метод генетики человека— близнецевый м е т о д
. Однояйцовые близнецы (ОБ) развиваются из одной яйцеклетки, оплодотворённой одним спермием; поэтому набор генов (генотип) у ОБ идентичен. Разнояйцевые близнецы (РБ) развиваются из двух и более яйцеклеток, оплодотворённых разными спермиями; поэтому их генотипы различаются
8-09-2015, 21:04