Сучасну цитологію на основі експериментальних досліджень застосовують при цитодіагностиці хвороб, цитоімунологічних пробах, у тканевоспецифічних регуляціях розмноження клітин, реактивних змін під впливом факторів зовнішнього середовища тощо.
Науково-технічний прогрес, успіхи розвитку методів дослідження дали змогу визначити ультраструктурну організацію клітини та неклітинних структур, зрозуміти процеси диференціювання, регенерації, передачі спадкових ознак тощо.
4. Біологія клітин: органели, їх будова і функції.
Серед живих організмів існує два типи організації клітин: прокаріотичні та еукаріотичні. До прокаріотичних відносять клітини бактерій та синьозелених водоростей, їх називають доядерними клітинами, вони обмежені плазматичною мембраною, зовні від плазматичної мембрани знаходиться клітинна стінка, яка є похідним клітинної активності. Прокаріотичні клітини не мають морфологічного ядра, але містять нуклеоідну зону, заповнену ДНК, а в матриксі цитоплазми прокаріотів розміщуються рибосоми.
Еука ріотичні клітини належать до вищого типу, їх обов'язковою структурою є: цитоплазма, клітинна оболонка (плазмолема) та клітинне ядро, яке відокремлене від цитоплазми ядерною мембраною. У клітинах еукаріотичного типу в цитоплазмі є спеціальні структури — органели, які виконують окремі специфічні функції. До органел відносять мембранні та немембранні структури (рис. 1).
У тварин найменшою одиницею живого є клітина еукаріотичного типу .
Узагальнюючи уяву про будову клітин, як одиниць живого, їх розмноження і ролі у формуванні багатоклітинних організмів, клітина являє собою елементарну саморегулюючу живу систему, основу будови, розвитку та життєдіяльності усіх тваринних і рослинних організмів.
Виходячи із положення клітинної теорії Т.Шванна, клітина є частина цілісного організму, найменша його одиниця, їй притаманні усі властивості, що відповідають визначенню поняття «живого»; клітини рослин і тварин мають однаковий загальний план будови і подібність загально-клітинних функцій для підтримання їх життя та розмноження, яке відбувається шляхом поділу. У багатоклітинному організмі клітини мають спеціалізацію будови і виконують різні функції. Однак вони підпорядковані цілому, входять до складу тканин, органів, систем органів, пов'язані міжклітинними та нейрогуморальними формами регуляції. Величезна різноманітність живої матерії та індивідуальність, що притаманні кожному організмові, різноманітність форм та функцій клітин у багатоклітинному організмі пов'язані з білками та нуклеїновими кислотами. Нуклеїнові кислоти забезпечують передачу спадкової інформації, відіграють велику роль у процесах біосинтезу білків. Білки беруть безпосередню участь у всіх реакціях, що відбуваються в клітинах та в організмі, характеризуються високим ступенем специфічності. Білкову природу мають усі ферменти, антитіла, деякі гормони тощо. До складу живої матерії входять вода, вуглеводи, ліпіди, вітаміни, мінеральні солі. Щодо фізико-хімічного стану клітина являє собою колоїдну систему, залежно від функціонального стану здатну переходити із рідкого в гелепо-дібний та в зворотній стан. Детальна інформація про фізико-хімічні особливості речовин, що входять до складу живої системи, вивчає наука біохімія.
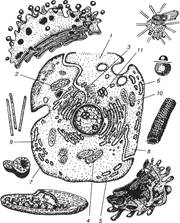
рис.1.Загальна схема будови клітини.
1 — плазмолема; 2 – комплексГольджі; 3 —цитоплазма; 4 — мітохондрія;
5 – ендоплазматичнасітка; 6 - рибосома; 7- лізосома; 8 - мікротрубочка;'
9 —мікрофіламенти; 10 - ядро; 11- центросома.
Структурні компоненти клітин.
Цитоплазма (cytoplasma), від навколишнього середовища відокремлена пла-змолемою; до неї належать гіалоплазма, органели, включення.
Плазмолема є різновидом мембран клітини. Це тонкі (6—10 нм) пласти ліпо- протеїдної природи, до складу яких входять 40% ліпідів, близько 60% білків, значна частина клітинних мембран містить 5—10% вуглеводів. Серед ліпідів знаходяться органічні речовини, які є гідрофобними — погано розчиняються у воді та характеризуються значною розчинністю в органічних розчинниках та жирах — ліпофільні. Серед ліпідів, що входять до складу клітинних мембран, знаходяться фосфоліпіди, сфінгомієліни, холестерин. Особливістю ліпідних мембран є поділ молекул на дві функціонально різні частини: гідрофобні неполярні, що не несуть зарядів, «хвости», складаються із жирних кислот, та гідрофільні, заряджені полярні «головки». Це визначає здатність ліпідів довільно утворювати двошарові (біліпідні) мембранні структури товщиною 5—7 нм. Клітинні мембрани можуть значно різнитися між собою за ліпідним складом та білковими молекулами. Багато мембранних білків складаються з двох частин, з ділянок багатих полярними (що несуть заряд) амінокислотами, та ділянок, збагачених неполярними амінокислотами. Такі білки в ліпідних шарах мембран розміщуються так, що їх неполярні ділянки ніби занурені в «жирну» частину мембрани, де знаходяться гідрофобні ділянки ліпідів. Полярна (гідрофільна) частина цих білків взаємодіє з головками ліпідів і оберальних білків, існують білки частково вмонтовані у мембрану — напівінтегральні та примембранні, вмонтовані в біліпідний шар. За біологічним значенням білки мембран поділяються на білки-ферменти, переносники, рецепторні та структурні.
Вуглеводи мембран входять до складу не у вільному стані, а у зв'язку з молекулами ліпідів або білків. Такі речовини називають відповідно гліколіпідами та глікопротеїдами. На поверхні плазмолеми вони формують надмембранну зону — глікокалікс, товщиною 3— 4 нм. За його участю здійснюється взаєморозпізнання клітин та взаємодія їх між собою. Всі мембрани є бар'єрними структурами.
Плазмолема виконує функції розмежування цитоплазми із зовнішнім середовищем, рецепції та транспорту різних речовин, серед яких важливе місце належить забезпеченню оптимального рівня асиметрії концентрації іонів натрію і калію у клітині та за її межами, а також забезпечує взаєморозпізнавання і взаємодію клітин з утворенням міжклітинних контактів, формування структури клітинної поверхні, рецепцію сигналізації з боку зовнішнього середовища.
Транспортна функція плазмолеми здійснюється шляхом дифузії пасивного перенесення різних речовин — води, іонів, деяких низькомолекулярних сполук. Інші речовини проникають через мембрану шляхом активного перенесення проти градієнта концентрації за рахунок розщеплення АТФ. Так транспортуються цукри, амінокислоти. Ці процеси відбуваються за участю спеціальних білків — переносників. Транспорт із зовнішнього середовища в клітину називають ендоцитозом, за межі клітини — екзоцитозом. Великі молекули та їх агрегати проникають у клітину шляхом фагоцитозу, який був вперше описаний І.І. Меч-никовим.
Поглинання частинок рідини називають піноцитозом. Загальним для цих процесів є те, що поглинання на поверхні плазмолеми відбувається шляхом оточення речовини ділянкою плазматичної мембрани у вигляді вакуолі, яка переміщується у цитоплазму. Поглинуті частинки розщеплюються за допомогою ферментів і їхні складові засвоюються клітиною. Таким чином, поглинуті речовини у середині мембранних вакуолей, утворених із елементів плазмолеми, підлягають внутрішньому клітинному травленню. Процес, при якому поглинені частинки в оточенні мембрани проходять через цитоплазму і виводяться без змін за межі клітини називають цитопемпсисом.
Плазмолема бере участь у виведенні речовин із клітини (екзоцитоз). У цьому випадку внутрішньоклітинні продукти (білки, мукополісахариди, жирові краплі тощо) містяться у вакуолі або міхурці і відмежовані від гіалоплазми мембраною наближаються до плазмолеми. У місцях контакту плазмолема та мембрана вакуолі зливаються і вміст вакуолі виходить за межі клітини.
Екзоцитоз поділяють на такі різновиди: секрецію — виділення клітиною продуктів її синтетичної діяльності; екскреція — виділення шкідливих продуктів метаболізму; клазматоз — видалення за межі клітин окремих її структурних компонентів.
Ендоцитоз та екзоцитоз відбуваються за участю пов'язаних з плазмолемою системи фібрилярних компонентів цитоплазми — мікротрубочок та скорочувальних мікрофіламентів, вони з'єднуються у певних ділянках плазмолеми і утворюють кортикальний шар.
Рецепторні функції плазмолеми пов'язані з локалізацією на ній спеціальних структур, які беруть участь в специфічному пізнанні хімічних та фізичних факторів. Клітинна поверхня має велику кількість рецепторів, що визначають можливість специфічних реакцій. Існують рецептори до біологічно активних речовин — гормонів, медіаторів, антигенів тощо. Складні процеси рецепції є основою взаєморозпізнання клітин, важливою і необхідною умовою існування багатоклітинних організмів.
Міжклітинні контакти
Сполучення між клітинами у складі тканин та органів багатоклітинних організмів утворені складними спеціальними структурами, які називають міжклітинними контактами. Розрізняють такі типи міжклітинних контактів. Просте міжклітинне сполучення — плазмолеми сусідніх клітин наближаються одна до одної на відстань 15—20 нм. При цьому відбувається взаємодія шарів глікокалікса. Розрізняють також щільне з'єднання за типом замка, при цьому шари двох плазмолем наближені максимально — ділянки плазмолем двох сусідніх клітин ніби зливаються. Ця область непроникна для молекул та іонів. Тип з'єднання плазмолем називають десмосомою. Вона являє собою площу діаметром до 0,5 мкм, інколи має вигляд шарів; між мембранами знаходиться зона з високою електронною щільністю; зміцнення зв'язку між клітинами досягається за допомогою фібрилярних структур цитоплазми та кортикального шару плазмолеми. Спостерігаються з'єднання типу напівдесмосом, якщо десмосома складається лише з однієї пластинки прикріплення.
Щільний замикаючий контакт характерний для апікальної поверхні клітин каймистого епітелію, а також ендотелію, мезотелію. Такий контакт характеризується максимальним зближенням плазматичних мембран сусідніх клітин, проміжок ущільнюється за рахунок анастомозуючих фібрил та іонів кальцію. При цьому зовнішні гідрофільні шари і глікокалікс суміжних плазмолем тісно зливаються в один суцільний шар завтовшки 2—3 нм. У серцевих м'язах зустрічаються щілинні контакти (нексуси), що забезпечують безпосередній обмін речовин між сусідніми клітинами. Так відбувається перенесення іонів та дрібних молекул.
Синаптичні з'єднання характерні для нервової тканини, або між нервовою клітиною і м'язом, у ділянці якого відбувається передача імпульсу.
Синапс — спеціалізований контакт, що забезпечує передачу нервового збудження. До його складу входять: ділянка плазмолеми пресинаптичної мембрани відростка нервової клітини, з якої відходить імпульс, ділянка плазмолеми постсинаптичної мембрани клітини, яка сприймає сигнал. До сигналу входить синаптична щілина, що розмежовує пресинаптичну та постсинаптичну мембрани і, заповнені нейромедіатором, синаптичні пухирці. Синапси забезпечують однобічну передачу інформації від клітини до клітини завдяки медіатору.
Цитоплазма.
До складу цитоплазми (Cytoplasma) входять 1) гіалоплазма, 2) органели та 3) включення.
1) Гіалоплазма — це прозора основна плазма, або матрикс цитоплазми , що позначає її внутрішнє середовище. У електронному мікроскопі матрикс цитоплазми має вигляд гомогенної та тонкозернистої речовини з низькою електронною щільністю. Гіалоплазма являє собою складну колоїдну систему і містить різні біополімери: білки, нуклеїнові кислоти, полісахариди тощо. Ця система здатна переходити із рідкого стану в гель і навпаки. Окремі зони гіалоплазми можуть змінювати свій агрегатний стан залежно від умов або функціонального завдання. Деякі молекули білків — тубулінів можуть бути диспергійовані в гіалоплазмі, але в певні моменти вони починають збиратися і утворювати мікротру-бочки. Таким же чином, у гіалоплазмі можуть виникати і розпадатися різноманітні фібрилярні, нитчасті комплекси білкових молекул. До ферментів матрикса відносять ферменти гліколізу, метаболізму цукрів, азотистих сполук, амінокислот, ліпідів та інших важливих сполук. У гіалоплазмі відбувається постійний рух іонів до плазматичної мембрани і від неї, до мітохондрій, ядра, вакуолі. Гіалоплазма є зоною переміщень молекул АТФ. В гіалоплазмі містяться органели та включення.
2) Органели — постійні мікроскопічні та ультраструктурні утворення, що виконують життєвоважливі функції клітин. Серед органел розрізняють мембранні органели — мітохондрії, ендоплазматичну сітку, лізосоми, пероксисоми, комплекс Гольджі; до немембранних органел відносяться — рибосоми, мікрофіла-менти, мікротрубочки, центросома. Всі вони належать до органел загального призначення. Поряд з цим в деяких клітинах існують спеціальні органели — тонофібрили епітеліальних клітин, міофібрили міоцитів та м'язових волокон, нейрофібрили нервових клітин.
Мембранні органели
Мітохондрії. Термін «мітохондрія» введено Бендою у 1897р. В світловому мікроскопі мітохондрії мають вигляд круглих зерен та коротких паличок. При розгляді під електронним мікроскопом кожна мітохондрія має овальну або видовжену форму. У них розрізняють зовнішню гладеньку та внутрішню мембрани, від останньої всередину мітохондрії відходять вирости — кристи. Проміжки між кристами заповнені матриксом — електронно-щільною речовиною, у якій виявляють ниткоподібні утворення товщиною 2—3 мм та гранули товщиною 15—20 мм. Ниткоподібні утворення матрикса являють собою молекули ДНК і РНК, а гранули — мітохондріальні рибосоми. У матриксі і у внутрішній мембрані містяться білки — ферменти, що забезпечують синтез АТФ шляхом окисного фосфорилювання аденозиндифосфату. Середній розмір сильно варіює і становить понад 20% загального об'єму цитоплазми і містить до 35% загальної кількості білка в клітині. Мітохондрії концентруються у тих місцях цитоплазми, де є потреба в АТФ. Основною функцією їх є синтез АТФ, який відбувається в результаті процесів окислення органічних субстратів і фосфорилювання АДФ.
Мітохондрії також беруть участь у регуляції обміну води, депонування іонів кальцію, продукції попередників стероїдних гормонів. Мітохондрії здатні рухатися; їх кількість збільшується шляхом поділу, або брунькуванням початкових мітохондрій. Мітохондрії чутливі до різних зовнішніх впливів: голодування, дії рентгенівських променів, наркотиків тощо.
Ендоплазматична сітка. Її вперше описав К.Портер у 1945 р. Вона являє собою субмікроскопічну органелу, яка утворює внутрішньоцитоплазматичну циркуляційну систему. Вона є замкненою сукупністю канальців, мішечків та цистерн, утворених безперервною біомембраною. Розрізняють два типи її — гранулярну та агранулярну. Ширина канальців гранулярної сітки від 20 до 1000 нм, з боку гіалоплазми її мембрани покриті рибосомами. Останні беруть участь у синтезі білків на «експорт» і білків-ферментів, а також білків, що нагромаджуються в цистернах ендоплазматичної сітки та здатні транспортуватися у вакуолі комплексу Гольджі, де перетворюються і входять до складу лізосом або секреторних гранул, утримання яких залишається ізольованим від гіалоплазми мембраною. У ряді випадків у канальцях або вакуолях гранулярної ендоплазматичної сітки відбувається модифікація білків, зв'язування їх з цукрами і утворення секреторних гранул. Гранулярна ендоплазматична сітка здатна синтезувати мембранні інтегральні білки, які вмонтовуються в товщу мембран.
На відміну від гранулярної ендоплазматичної сітки на мембранах агранулярної ендоплазматичної сітки відсутні рибосоми. Діаметр її канальців і міхурців 50— 100 нм. Функція гладкої ендоплазматичної сітки пов'язана з метаболізмом ліпідів, синтезом глікогену, депонуванням іонів кальцію, дезактивацією отруйних речовин, що особливо характерно для гепатоцитів. Мембрана ендоплазматичної сітки безпосередньо контактує з плазмолемою клітин та мембранами.
Лізосоми — мембранні органели. Вони являють собою кулясті структури розміром 0,2— 0,4 мкм, містять понад 60 гідролітичних ферментів, здатних до розщеплення біополімерів різної хімічної природи. Із ферментів лізосоми містять протеїнази, нуклеази, глюкозидази, фосфатази, ліпази. Ферменти локалізуються у лізосомах і утримуються у них за допомогою ліпопротеїнової мембрани, яка обмежує і відокремлює їх вміст від зовнішніх субстратів. Основна фізіологічна функція лізосом — внутрішньоклітинне травлення в зв'язку з процесами фагоцитозу, піноцитозу тощо. Залежно від ультраструктурних та функціональних особливостей лізосоми поділяють на первинні, їх ферменти знаходяться у неактивному стані; вторинні, або фагосоми (активовані ферменти в них безпосередньо контактують з розщеплюваними біополімерами), а також залишкові тільця, оточені біомембраною нерозщеплені залишки. Поряд з цим лізосоми можуть брати участь у розщепленні власних макромолекулярних комплексів клітин — аутофагоцитозу.
Пероксисоми — субмікроскопічні мембранні органели. Відіграють вирішальну роль у процесах детоксикації клітини. Пероксисоми — тільця розміром 0,3-1,5 мкм, обмежені мембраною, містять гранулярний матрикс, в центрі якого розміщуються кристолоїдноподібні структури із фібрил і трубок. Ферментні системи пероксисом (каталаза) спрямовані на утилізацію хімічно активного атомарного кисню, а також забезпечують розщеплення етилового спирту, сечової кислоти, регуляцію обміну ліпідів.
Комплекс Гольджі. Перші відомості про комплекс Гольджі належить К.Гольджі, який в 1898р. описав цю органелу у складі нервових клітин. Під світловим мікроскопом комплекс Гольджі має вигляд сітчастого утворення. Під електронним мікроскопом він являє собою мембранні структури, що мають вигляд цистерн товщиною до 25 нм, сплющених у центральній частині і розширених на периферії. Окрему зону скупчення цих мембран називають діктіосомою. В проміжках між окремими цистернами знаходяться тонкі прошарки гіалоплазми. У комплексі Гольджі завершується процес формування продуктів синтетичної діяльності клітини — її кінцеве глікозування. Комплекс Гольджі бере участь у сегрегації і нагромадженні продуктів, синтезованих в цитоплазматичному ре-тикулумі, у їх хімічній трансформації. У цистернах комплексу Гольджі синтезуються полісахариди, які комплексуються з білками, що сприяє утворенню мукопротеїдів, а також виведенню готових секретів за межі клітини. Крім того, комплекс Гольджі забезпечує формування клітинних лізосом.
Не мембранні органели
Рибосоми. За розміром рибосоми становлять 25x20x20 нм. До їх складу входять велика і мала субодиниці рибосомної РНК та білок.
Для стабілізації структури рибосом необхідні іони магнію. Кожна із субоди-ниць побудована з нуклеопротеїдного тяжа, де РНК взаємодіє з різними білками і утворює тіло рибосоми, у яких амінокислоти сполучаються, тобто синтезуються білкові молекули. Кілька рибосом, з'єднаних спільною ниткою інформаційної РНК, називають полісомами. Останні полісоми пов'язані з мембранами ендоплазматичної сітки, синтезують білки для виведення за межі клітини. Ступінь інтенсивності синтетичної діяльності вільних рибосом менший, а утворені білки використовуються для внутрішніх потреб клітини.
Мікрофіламенти. Це тонкі волоконця діаметром до 5 нм, побудовані з різних білків — актину, міозину та ін. Вони розміщені переважно в кортикальній зоні клітини у складі її цитоплазматичних виростів і виконують роль цитоскелету, а також скоротливого апарату клітини.
Мікротрубочки являють собою прямі, довгі порожнисті білкової природи циліндри. їх зовнішній діаметр досягає 25 нм, а внутрішній просвіт — 15 нм; містять глобулярний білок тубулін. Мікротрубочки входять до складу центросоми та базальних тілець, а також є основними структурними елементами війок та джгутиків. Мікротрубочки забезпечують рухливість клітинних органел, а також беруть участь в утворенні цитоскелету.
Центросома. Описана В.Флемінгом у 1875р. Міститься вона у клітинах тварин, за винятком яйцеклітин. Центросома складається з двох центріолей, оточених центросферою, що являє собою позбавлену органел гіалоплазму, яку радіально пронизують мікрофіламенти і мікротрубочки. Основою будови центріолей є дев'ять триплетів паралельно розміщених мікротрубочок, які формують циліндр 200x500 нм. Крім мікротрубочок, до складу центріолі входять так звані «ручки», за їх допомогою триплети пов'язані між собою. У складі «ручок» міститься білок тубулін, що має АТФ-азну активність і якому належить важлива роль у механізмі рухових функції центріолей. Довгі осі обох центріолей розміщені у взаємно перпендикулярних площинах. Перед поділом клітини центріолі подвоюються (настає дупликація) з подальшим розходженням кожної новоутвореної пари до полюсів клітини. Дві розміщені поряд центріолі
8-09-2015, 22:51