Со времени открытия архебактерий в 1977 г. количество относящихся к ним организмов и объем знаний о последних возрастают стремительно. К архебактериям относят 5 групп прокариотных организмов. Две из них — метанобразующие бактерии и экстремальные галофилы — известны давно. Более 10 лет известны и представители двух других групп: термоацидофильные серные аэробные бактерии и микоплазмоподобная форма Thermoplasma acidophilum . Последняя группа, обнаруженная в начале 80-х гг., — анаэробные формы, метаболизм которых связан с молекулярной серой.
По сравнению с эубактериями число известных архебактерий мало, поэтому при описании мира прокариот во II разделе внимание было сосредоточено на эубактериях. Настоящий раздел посвящен общей характеристике (гл. 17) и отдельным группам (гл. 18) архебактерий.
ГЛАВА 17. АРХЕБАКТЕРИИ: ОБЩАЯ ХАРАКТЕРИСТИКА
Формы клеток архебактерий в целом сходны с таковыми эубактерий (см. рис. 3). Среди них есть кокки, палочки, извитые клетки и виды, характеризующиеся слабым ветвлением. Особенность архебактерий — отсутствие сложных многоклеточных форм, мицелиальных и трихомных, достаточно хорошо представленных у грамположительных и грамотрицательных эубактерий.
По тонкому строению клетки, выявляемому с помощью электронного микроскопа, архебактерии принципиально не отличаются от эубактерий и ближе к грамположительной их ветви. Прокариотная организация архебактерий проявляется в отсутствии у них ядра и характерных для эукариот органелл, окруженных мембраной. Хромосомная ДНК организована в виде нуклеоида, т. е. расположена непосредственно в цитоплазме и имеет вид электроннопрозрачной зоны, заполненной нитями ДНК.
От внешней среды клетки отделены клеточной стенкой (исключение составляет Thermoplasma acidophilum ). У одних видов она выглядит как толстый гомогенный слой, у других — тонкий, структурированный. У некоторых нитчатых форм поверх клеточной стенки расположен чехол, объединяющий несколько клеток. Многие виды имеют жгутики и ворсинки эубактериального типа. В цитоплазме некоторых архебактерий обнаружены газовые вакуоли и запасное вещество гликоген, присущие многим эубактериям.
У архебактерий описаны разные способы размножения: равновеликое бинарное деление, почкование, фрагментация. Все они имеются и у эубактерий.
Хотя клетки архебактерий структурно относятся к прокариотному типу, многие макромолекулы, входящие в их состав (липиды, полисахариды, белки), уникальны и 'не 'найдены ни у эубактерий, ни у эукариот. Одно из существенных отличий архебактерий связано с химическим составом клеточных стенок, в которых не обнаружен характерный для эубактерий пептидогликан (см. рис. 6). В составе последнего в качестве обязательного компонента присутствует N -ацетилмурамовая кислота. Отсюда и часто употребляющееся его название муреин69 . Вместо него у ряда архебактерий из группы метанобразующих найден другой пептидогликан, получивший название псевдомуреина (рис. 101). Его гликановый остов построен из N -ацетилглюкозамина и N -ацетилталозаминуроновой кислоты, а пептидные фрагменты — только из L-аминокислот. Последовательность аминокислот пептидного хвоста псевдомуреина отличается от таковой эубактериального .пептидогликана. Кроме того, аминосахара гликановой цепи псевдомуреина соединены не с помощью b-1,4-, а через (b-1,3-гликозидные связи. Отсутствие у архебактерий муреина привело к их устойчивости к пенициллину и некоторым другим антибиотикам, ингибирующим синтез пептидогликана эубактерий. Под электронным микроскопом клеточные стенки такого типа, окрашивающиеся положительно по Граму, выглядят как однородный слой толщиной 15 — 40 нм, ничем морфологически не отличающийся от клеточной стенки грамположительных эубактерий.
69 От лат. murus — стенка.
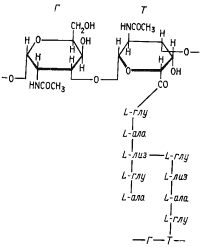 |
| Рис. 101. Структура псевдомуреина архебактерий: Г — N -ацетилглюкозамин; Т — N -ацетилталозаминуроновая кислота; глу — глутаминовая кислота; ала — аланин; лиз — лизин |
Описаны метанобразующие архебактерии с очень толстой (до 500 нм) аморфной клеточной стенкой, дающей положительную реакцию по Граму, построенной исключительно из кислого гетерополисахарида, в составе которого обнаружены галактозамин, нейтральные сахара и уроновые кислоты. Наличие у этих бактерий положительного окрашивания по Граму может служить указанием на то, что оно определяется не химическим составом клеточной стенки, а ее строением70 .
70 Вероятно, все гораздо сложнее. Описаны эубактерии, имеющие типичную грамположительную клеточную стенку, но окрашивающиеся по Граму отрицательно, и наоборот.
Наконец, у галобактерий, ацидофильно-термофильных архебактерий и большинства метанобразующих бактерий клеточная стенка построена из белка. В некоторых случаях в следовых количествах обнаружены аминосахара. Под электронным микроскопом клеточная стенка выглядит обычно как ряд регулярно расположенных белковых субъединиц. Все архебактерии с клеточной стенкой белковой природы грамотрицательны.
Другое уникальное свойство архебактерий касается состава их мембранных липидов. У них не найдены обычные для эубактерий эфиры глицерина и жирных кислот (см. рис. 14), но присутствуют эфиры, образованные путем конденсации глицерина с терпеноидными спиртами: диэфир состоит из глицерина, связанного простыми эфирными связями с двумя молекулами C20 -спирта фитанола; тетраэфир образован двумя остатками глицерина, соединенными двумя одинаковыми парами C40 -бифитанильных цепей (рис. 102, 1, 2 ). Молекула тетраэфира, таким образом, структурно эквивалентна двум молекулам диэфира. Бифитанильные цепи тетраэфиров могут быть ациклическими или содержать от 1 до 4 пятичленных колец (рис. 102, 3 ).
В мембранах архебактерий присутствуют до 80–90% полярных фосфо- и гликолипидов, образованных на основе ди- и тетраэфиров. Экстремально галофильные архебактерии содержат диэфиры в качестве единственных мембранных гликолипидов. В мембранах ацидотермофильных архебактерий почти все гликолипиды представлены тетраэфирами. Метанобразующие бактерии содержат ди- и тетраэфиры, соотношение их в мембранах зависит от вида. Наличие пятичленных колец в бифитаниловых цепях характерно для термоацидофильных архебактерий, и это понятно, так как эти химические структуры способствуют стабилизации мембраны, снижая ее текучесть и обеспечивая функционирование при высоких температурах. При повышении температуры количество цепей, содержащих циклические группировки, возрастает, ациклических — снижается. Кроме того, в зависимости от температуры культивирования меняется число колец в цепи.
Помимо полярных липидов архебактерии содержат нейтральные липиды, основными из которых являются изопреноидные углеводороды, насыщенные или содержащие двойные связи, — производные C15 -C30 -изопреноидных скелетов. Особенно распространены у архебактерий C30 -изопреноиды. Больше всего углеводородов содержится у метанобразующих бактерий, меньше — у галофилов и термоацидофилов.
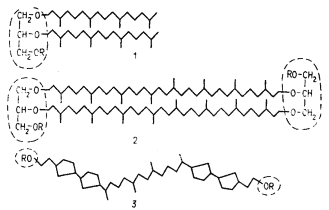 |
| Рис. 102. Липиды архебактерий: 1 — фитаниловый диэфир глицерина; 2 — бифитаниловый тетраэфир диглицерина; 3 — изопреноид из полярных липидов, содержащий пятичленные циклические группировки; обведены полярные области; R — остатки фосфорной кислоты или сахара |
Доминирование в мембране архебактерий липидов, образованных на основе ди- и тетраэфиров, поставило вопрос о принципиальной ее организации. По современным представлениям, у всех эубактерий и эукариот основу элементарной (липопротеиновой) мембраны составляет липидный бислой (см. рис. 15). Диэфиры архебактерий способны образовывать элементарные мембраны, состоящие из двух ориентированных слоев липидных молекул. Молекулы тетраэфира имеют длину порядка 5–7,5 нм. Толщина мембраны архебактерий примерно 7 нм. Такая мембрана не может быть организована из двух слоев тетраэфирных молекул. Очевидно, что в данном случае она представляет собой липидный монослой (рис. 103). Монослойные липидные мембраны обладают, очевидно, повышенной жесткостью по сравнению с бислойными. Обнаружение липопротеиновой мембраны, в основе которой лежит иной принцип организации, приводит к отказу еще от одной догмы — универсальности элементарной мембраны с бислойным липидным строением.
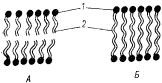 |
| Риc. 103. Схема бислойной и монослойной мембран архебактерий, образованных соответственно ди- (А) и тетраэфирами (Б) глицерина: 1 — молекула глицерина; 2 — углеводородные цепи разной длины (по Воробьевой, 1987) |
Существенные отличия выявлены у архебактерий в строении генома, аппаратов репликации, транскрипции и трансляции. Прежде всего исследователи обратили внимание на то, что именно в группе архебактерий наименьший геном среди свободноживующих форм прокариот: у Thermoplasma acidophilum — 0,8x109 Да, изученных метанобразующих бактерий — порядка 1x109 Да (для сравнения молекулярная масса генома Е. coli — 2,5x109 Да). Однако у галобактерий величина генома оказалась больше, чем у Е. coli. Особенность генома архебактерий — наличие многократно повторяющихся нуклеотидных последовательностей, а в генах, кодирующих белки, тРНК и рРНК, — интронов, что характерно для организации генетического материала эукариот. У некоторых архебактерий обнаружены основные гистоноподобные белки, связанные с ДНК. Функция их предположительно заключается в обеспечении определенной упаковки ДНК в нуклеоиде.
Помимо хромосомной ДНК в клетках архебактерий обнаружены типичные для эубактерий фаги, плазмиды, мигрирующие элементы.
Существование механизмов переноса генетической информации с помощью фагов и плазмид позволяет предполагать, что архебактерии должны каким-то образом защищать собственный генетический материал от чужеродного. У эубактерий эта проблема решена с помощью системы рестрикции-модификации. У эукариот такой системы нет, они выработали иные механизмы генетической изоляции. Найдено, что архебактерии обладают системой рестрикции-модификации, аналогичной эубактериальной.
Информация об аппарате репликации архебактерий в основном ограничивается данными о выделенной из ограниченного числа видов ДНК-зависимой ДНК-полимеразе, по некоторым свойствам близкой к эукариотному типу. Генетический код архебактерий такой же, как у других организмов.
ДНК-зависимая РНК-полимераза архебактерий сочетает свойства, характерные для эукариот и эубактерий. У всех изученных представителей архебактерий РНК-полимераза одной формы, осуществляющая, как и в случае эубактерий, транскрипцию всех генов. (У эукариот, например дрожжей, существуют 3 формы РНК-полимеразы, различающиеся функционально, компонентным составом, чувствительностью к ингибиторам). Фермент архебактерий отличается структурной сложностью, в его состав входят от 5 до 11 отдельных субъединиц. (РНК-полимеразы эубактерий состоят из 4 — 8 компонентов, а эукариот — 10 — 14 субъединиц). Характерным для РНК-полимераз всех эубактерий является их чувствительность к антибиотикам, специфически ингибирующим инициацию (рифампицин) и элонгацию (стрептолидигин 1) транскрипции. Архебактериальная и все РНК-полимеразы эукариот не чувствительны к этим антибиотикам.
Недавно у архебактерий описан известный только у эукариот процессинг: вырезание из первичного продукта транскрипции определенных нуклеотидных участков, укорачивание и образование зрелых молекул РНК.
Процесс трансляции у архебактерий происходит по тому же принципиальному пути, что и у других организмов, но обнаружены многочисленные особенности в организации трансляционного аппарата. Рибосомы архебактерий сочетают свойства, присущие эубактериям и эукариотам: по размерам они схожи с рибосомами эубактерий (имеют константу седиментации 70S, а их субъединицы — 30S и 50S), по форме ближе к 80S рибосомам эукариот.
Состав рибосомальных РНК архебактерий типично эубактериальный (5S, 16S и 23S рРНК), но их первичные структуры отличны от эубактериальных и эукариотных. Изучение нуклеотидных последовательностей 16S (18S) рРНК разных представителей живого мира и привело к выявлению среди прокариот группы архебактерий. Значения коэффициента сходства (SAB ), отделяющие рРНК эубактерий, архебактерий и эукариот друг от друга, лежат в области 0,1 (SAB , равный 1, соответствует полной гомологии нуклеотидных последовательностей; SAB порядка 0,02 — уровень случайного совпадения).
Архебактериальные 5S рРНК по нуклеотидной последовательности также заметно отличаются от соответствующих рРНК эубактерий и эукариот. Вторичные структуры этих РНК у различных представителей архебактерий проявляют наличие эубактериальных, эукариотных и уникальных черт в разных соотношениях и по своему разнообразию охватывают широкий спектр структур от типично эубактериальной до типично эукариотной.
Количество рибосомальных белков у архебактерий больше, чем у эубактерий, но меньше, чем у эукариот. Получены данные, свидетельствующие об уникальности первичной структуры ряда рибосомальных белков архебактерий. В то же время наиболее интенсивно изучаемый рибосомальный белок А архебактерий по аминокислотной последовательности сходен с соответствующим белком эукариот. На 70S архебактериальной рибосоме отсутствуют места связывания ингибиторов, специфичных для 70S рибосом эубактерий (хлорамфеникол, стрептомицин), но есть места для связывания ингибиторов, специфических в отношении 80S рибосом эукариот.
Для тРНК архебактерий характерно сходство с остальными организмами по общим принципам организации, но значительные различия по многим частным деталям. Это проявляется, например, в специфической модификации ряда нуклеотидов в молекуле архебактериальной тРНК, происходящей после окончания транскрипции. Модификации могут быть уникальными для архебактерий, общими для эубактерий и эукариот, характерными только для эубактерий или только для эукариот. В целом тРНК архебактерий отличается от тРНК эубактерий и эукариот в такой же степени, как последние различаются между собой.
Несмотря на выявленные многочисленные различия в аппаратах транскрипции и трансляции между эубактериями и архебактериями, было показано, что архебактериальная ДНК, перенесенная с помощью плазмиды в клетку эубактерий, может считываться в ней, результатом чего является синтез функционально активных ферментов.
Метаболизм архебактерий. В группе архебактерий известны организмы с хемоорганогетеротрофным, хемолитоавтотрофным, хемолитогетеротрофным и фотогетеротрофным типом питания. Источником углерода для разных представителей могут служить сахара, аминокислоты, органические кислоты, C1 -соединения и CO2 . У архебактерий, метаболизм которых связан с молекулярной серой, автотрофная фиксация CO2 происходит по восстановительному ЦТК, функционирующему также в группе зеленых серобактерий (см. рис. 76); виды, способные расти на среде с CO2 + H2 , фиксируют углекислоту по недавно открытому нециклическому ацетил-КоА-пути, описанному у ацетогенных эубактерий (см. рис. 62). Центральным метаболитом этого пути является ацетил-КоА, который через фосфоенолпируват превращается в 3-ФГА и далее в молекулу фруктозо-6-фосфата. Таким образом, у архебактерий, способных к автотрофии, фиксация CO2 происходит по восстановительному пути карбоновых кислот в различных его модификациях. Ни в одном случае не обнаружено функционирование восстановительного пентозофосфатного цикла. В сравнении с последним и восстановительным ЦТК ацетил-КоА-путь фиксации CO2 более экономичен с точки зрения потребления АТФ, но требует большего количества восстановленных кофакторов71 .
Исследование других реакций конструктивного метаболизма архебактерий, как правило, обнаруживает пути, функционирующие у эубактерий. Так, глюконеогенез, начиная с пирувата, идет по тому же механизму, что у эубактерий. Ассимиляция аммония, синтез изопреноидных липидов, нуклеотидов происходит по обычным для эубактерий путям.
Катаболизирование архебактериями Сахаров происходит по путям, свойственным эубактериям: гликолиз, окислительный пентозофосфатный путь, ЦТК и путь Энтнера–Дудорова. Эти катаболические пути найдены не у всех представителей группы. У многих архебактерий, например, отсутствует гликолиз. Таким образом, анаболические и катаболические пути превращения углеродных соединений у архебактерий сходны с эубактериальными путями.
71 Для синтеза 1 молекулы триозы в восстановительном пентозофосфатном цикле требуется 9 молекул АТФ, в восстановительном ЦТК — 5, а в ацетил-КоА-пути — только 3.
Способы получения архебактериями энергии включает бесхлорофилльный фотосинтез, брожение, аэробное и анаэробное дыхание, при котором конечными акцепторами электронов могут быть CO2 и другие C1 -соединения, молекулярная сера, NO3 – , Fe3+ и Mo6+ . У организмов, получающих энергию с использованием электронного транспорта, в качестве электронпереносящих компонентов обнаружены ферредоксины, хиноны, цитохромы. Электронный транспорт сопряжен с трансмембранным переносом протонов. Механизм окислительного фосфорилирования архебактерий соответствует хемиосмотическому принципу и сходен с аналогичным механизмом эубактерий и митохондрий. В то же время следует подчеркнуть, что архебактериям свойственны типы энергетического метаболизма, не встречающиеся у эубактерий и эукариот. Это бесхлорофилльный фотосинтез и особый тип анаэробного дыхания, в процессе которого происходит образование метана.
Таблица 33. Свойства архебактерий (по Дуде, 1985)
| Признаки, характерные для | Признаки, уникальные для архебактерий | |
| эубактерий | эукариот | |
| Нуклеоид Жгутики Бактериофаги, плазмиды Газовые вакуоли Фимбрии Серное дыхание Типы автотрофной фиксации CO2 Азотфиксация Типы ферредоксинов и цитохромов Один из типов клеточных стенок (псевдомуреин) Система рестрикции-модификации Размер рибосом и рРНК Типы некоторых рибосомальных белков |
Повторяющиеся последовательности в хромосомной ДНК Гистоны, связанные с хромосомной ДНК Наличие интронов в генах Процессинг Родопсиновый белок Форма рибосом и строение некоторых рибосомальных белков Чувствительность к некоторым антибиотикам, угнетающим клетки эукариот |
Клеточные стенки Особые мембранные липиды Монослой липидов в ЦПМ некоторых видов Субъединичный состав РНК-полимеразы Специфические кофакторы Образование метана Тип фотосинтеза Особенности нуклеотидного состава 5S, 16S рРНК и тРНК Сложный компонентный состав рибосомальных белков Способность некоторых видов расти при температурах выше 100° |
Некоторые свойства архебактерий, уникальные, сближающие их с эубактериями или эукариотами, суммированы в табл. 33. Ряд свойств архебактерий нельзя отнести ни к одной из перечисленных групп. Они у разных подгрупп различны и охватывают диапазон от типично эубактериальных до типично эукариотных. Обращает на себя внимание определенная "эволюционная ограниченность" этой группы прокариот. В ней отсутствуют патогенные, паразитические формы. Способность использовать органические вещества у большинства ограничена простыми низкомолекулярными соединениями, что связывают с неспособностью синтезировать активные гидролитические ферменты. Клеточная организация архебактерий не обнаруживает той степени сложности, которая свойственна грамотрицательным эубактериям. Нет у архебактерий циклов развития, характерных для ряда эубактерий.
Для архебактерий как группы в целом характерна способность существовать в широком диапазоне условий внешней среды. Среди них есть строгие и факультативные анаэробы и облигатные аэробы, нейтрофилы и облигатные ацидофилы, экстремальные галофилы. В этой же группе наряду с мезофилами описаны экстремальные термофилы, имеющие оптимальную температуру роста свыше 100° (см. рис. 36). Именно к этой группе прокариот относятся бактерии, растущие при самых высоких температурах. К архебактериям предположительно относятся микроорганизмы, обнаруженные на дне океана на глубине около 2,5 км, где давление достигает 260 атм, а температура воды в зонах выходящих со дна "черных гейзеров" — 250 — 300°.
При попытке охватить группу архебактерий в целом на первый план выступают черты высокой степени неоднородности внутри самой группы. По многим признакам архебактерий проявляют гораздо больше отличий друг от друга, чем эубактерий и эукариоты. На основании сравнительного исследования нуклеотидных последовательностей 16S рРНК в группе архебактерий выявлены 2 ветви: одна объединяет метанобразующие и экстремально галофильные виды, другая — аэробные и анаэробные серозависимые термо- и термоацидофилы. Промежуточное положение занимает термоацидофильная микоплазма Thermoplasma acidophilum. Максимальные "эволюционные расстояния" между разными группами архебактерий, выраженные с помощью коэффициента SAB , достигают величины, равной 0,2. Такая же величина SAB характеризует и филогенетически далекие группы эубактерий. По молекулярной организации ветвь, включающая метанобразующие и экстремально галофильные архебактерий, ближе к эубактериям, а серозависимые архебактерий — к эукариотам.
Данные, суммированные в табл. 33, говорят об обособленности группы архебактерий, их особом эволюционном пути и
29-04-2015, 01:57