Живые организмы, все клетки и, в частности, нейроны обладают раздражимостью -способностью реагировать на различного рода воздействия. Реакции носят сложный, комплексный характер, но всегда сопровождаются изменением электрического заряда мембраны. Воздействия, вызывающие реакции, называются раздражителями, или стимулами. Хотя живые организмы состоят из клеток, реакции организма в целом не сводятся к реакциям отдельных клеток. Напомним универсальное правило: закономерности системы не могут быть сведены к закономерностям отдельных элементов.
Нервные клетки и некоторые ткани (мышечная, железистая) специально приспособлены к осуществлению быстрых реакций на раздражение. Такие клетки и ткани называются возбудимыми, а их реакция -возбудимостью. Клетки при возбуждении переходят из состояния физиологического покоя в состояние активности. В частности, нервная клетка генерирует один, или несколько электрических импульсов. Это связано с тем, что при возбуждении проницаемость мембраны для ионов резко увеличивается. Возникают ионные токи, что естественно приводит к изменению разности потенциалов на поверхности мембраны. Это изменение мембранного потенциала называется потенциалом действия. Генерируемые нервными клетками импульсы также называются спайками. Группы, тесно следующих друг за другом спайков, называются пачками или берстами. Амплитуды спайков относительно стабильны для каждого типа нейронов, поэтому часто говорят, что нейрон реагирует на раздражение по закону “все, или ничего”.
Нервное возбуждение - распространяющийся процесс. Возникнув в одном, или нескольких нейронах, оно передается другим клеткам. Проведение возбуждения обусловлено тем, что потенциал действия, возникнув в одной клетке, становится раздражителем для связанных с ней клеток. То же самое относится к другим возбудимым тканям, например, к сердечной мышце. Согласно наиболее распространенной в настоящее время точки зрения считается, что “сообщения” в нервной системе передаются в виде импульсов “все, или ничего” в форме частотного кода.
Обсудим природу мембранного потенциала. Мембрана, отделяющая цитоплазму нейрона от межклеточной среды, состоит в основном из липидов -жироподобных веществ. В нее встроены белковые молекулы. Некоторые молекулы пронизывают липидный слой насквозь, а другие погружены лишь до сердцевины (своеобразные ворсинки). Некоторые белки выполняют роль каналов, осуществляющих транспорт ионов через мембрану. Другие -служат рецепторами, позволяющими нейрону реагировать на поступающие от других нейронов потенциалы действия.
Механизмы переноса ионов через мембрану делятся на два класса: пассивный и активный транспорт. Пассивный транспорт не требует затрат энергии. Он определяется двумя факторами. Диффузия способствует выравниванию концентрации растворов. Носители зарядов -ионы перемещаются под действием электрического поля. При активном транспорте за счет химической энергии, освобождающейся при окислении аминокислот, может осуществляться перенос ионов как в области с более высокой их концентрацией, так и против электрических сил.
Закономерности пассивного транспорта опираются на некоторые положения электрохимии. Как уже говорилось, мембраны биологического происхождения обладают избирательной проницаемостью. Одни ионы достаточно легко диффундируют через них. Для других ионов мембраны практически непроницаемы. Рассмотрим на примере положение об электрохимическом равновесии. Пусть в сосуд ![]() с раствором
с раствором ![]() помещена полость
помещена полость ![]() , ограниченная мембраной, где также находится раствор
, ограниченная мембраной, где также находится раствор ![]() . Обозначим через
. Обозначим через ![]() и
и ![]() концентрации раствора в сосуде и в полости. Предположим, что мембрана проницаема для ионов
концентрации раствора в сосуде и в полости. Предположим, что мембрана проницаема для ионов ![]() и непроницаема для ионов
и непроницаема для ионов ![]() . Пусть концентрация раствора в сосуде ниже, чем в полости, т.е.
. Пусть концентрация раствора в сосуде ниже, чем в полости, т.е. ![]() . Тогда ионы калия будут переходить из полости в сосуд, поскольку диффузия стремится выровнять концентрации. В полости образуется относительный дефицит положительно заряженных ионов и она заряжается отрицательно. Точнее, отрицательный заряд сосредотачивается на внутренней поверхности мембраны. Наоборот, внешняя поверхность мембраны заряжается положительно. Электрическое поле направлено так, что диффузия положительных ионов
. Тогда ионы калия будут переходить из полости в сосуд, поскольку диффузия стремится выровнять концентрации. В полости образуется относительный дефицит положительно заряженных ионов и она заряжается отрицательно. Точнее, отрицательный заряд сосредотачивается на внутренней поверхности мембраны. Наоборот, внешняя поверхность мембраны заряжается положительно. Электрическое поле направлено так, что диффузия положительных ионов ![]() будет затруднена. в результате при некоторой разности потенциалов
будет затруднена. в результате при некоторой разности потенциалов ![]() между внутренней и внешней поверхностью мембраны диффузия ионов вообще прекратится. Величина
между внутренней и внешней поверхностью мембраны диффузия ионов вообще прекратится. Величина ![]() называется равновесным электрохимическим потенциалом. Она вычисляется по формуле Нернста:
называется равновесным электрохимическим потенциалом. Она вычисляется по формуле Нернста:
![]() , (1)
, (1)
где ![]() -универсальная газовая постоянная,
-универсальная газовая постоянная, ![]() -абсолютная температура,
-абсолютная температура, ![]() -число Фарадея,
-число Фарадея, ![]() -заряд иона.
-заряд иона.
Пусть ![]() текущее значение мембранного потенциала. привлечем некоторые положения теории электрических цепей. Мембрана является диэлектриком (изолятором) и поэтому обладает емкостью. Ток
текущее значение мембранного потенциала. привлечем некоторые положения теории электрических цепей. Мембрана является диэлектриком (изолятором) и поэтому обладает емкостью. Ток ![]() , текущий через емкость и напряжение
, текущий через емкость и напряжение ![]() (оно же мембранный потенциал) связаны соотношением:
(оно же мембранный потенциал) связаны соотношением: ![]() , где коэффициент пропорциональности
, где коэффициент пропорциональности ![]() называется емкостью. Отметим, что в некотором смысле ток, текущий через мембрану, является фиктивным. Он не сопровождается переносом зарядов. Согласно закону Нернста мембрана является источником напряжения
называется емкостью. Отметим, что в некотором смысле ток, текущий через мембрану, является фиктивным. Он не сопровождается переносом зарядов. Согласно закону Нернста мембрана является источником напряжения ![]() . Ток
. Ток ![]() , текущий через источник напряжения
, текущий через источник напряжения ![]() , связан с напряжением
, связан с напряжением ![]() на его полюсах формулой:
на его полюсах формулой: ![]() . По закону Кирхгофа сумма токов равна нулю:
. По закону Кирхгофа сумма токов равна нулю: ![]() . Таким образом, получаем дифференциальное уравнение:
. Таким образом, получаем дифференциальное уравнение:
![]() . Cu
¢
+gK
(u
-
EK
)
(2)
. Cu
¢
+gK
(u
-
EK
)
(2)
Проведем тривиальный анализ уравнения (2). Перепишем его в виде:
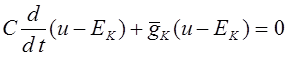 .
.
Отсюда получаем:
![]() .
.
Таким образом, по экспоненте мембранный потенциал ![]() при
при ![]() .
.
Сделаем ряд замечаний. При анализе уравнения (2) мы предполагали, что коэффициенты ![]() и
и ![]() не зависят от
не зависят от ![]() . Действительно для мембраны емкость практически постоянна. Однако, проводимость
. Действительно для мембраны емкость практически постоянна. Однако, проводимость ![]() есть функция от
есть функция от ![]() . Проведенный анализ справедлив лишь при малых отклонениях
. Проведенный анализ справедлив лишь при малых отклонениях ![]() от равновесного значения
от равновесного значения ![]() .
.
Пусть теперь дополнительно к ![]() в полости и в сосуде присутствует
в полости и в сосуде присутствует ![]() , концентрации которого в полости и в сосуде различны. Ионам
, концентрации которого в полости и в сосуде различны. Ионам ![]() отвечает свое равновесное значение
отвечает свое равновесное значение ![]() . Натриевый ток
. Натриевый ток ![]() связан с мембранным потенциалом
связан с мембранным потенциалом ![]() соотношением:
соотношением: ![]() . Привлекая снова закон Кирхгофа, получаем:
. Привлекая снова закон Кирхгофа, получаем:
![]()
![]() . . Cu
¢
+gNa
(u
-
ENa
)
+gK
(u
-
EK
) (3)
. . Cu
¢
+gNa
(u
-
ENa
)
+gK
(u
-
EK
) (3)
Теперь в силу (3) равновесное значение мембранного потенциала суть:
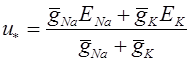 .
.
Одна из первых гипотез о природе мембранного потенциала была высказана в 1902 г. Бернштейном, который использовал положения электрохимии. Он предположил, что предположил, что концентрация ионов ![]() в клетке выше, чем во внеклеточной среде. Вследствие этого ионы
в клетке выше, чем во внеклеточной среде. Вследствие этого ионы ![]() диффундируют из клетки. Образуется их относительный дефицит положительных ионов внутри клетки и внутренняя поверхность мембраны заряжается отрицательно. Тем самым, потенциал покоя определяется равновесным электрохимическим потенциалом для ионов
диффундируют из клетки. Образуется их относительный дефицит положительных ионов внутри клетки и внутренняя поверхность мембраны заряжается отрицательно. Тем самым, потенциал покоя определяется равновесным электрохимическим потенциалом для ионов ![]() . Далее, Бернштейн предположил, что в жизни нейрона иногда наступает особый момент -состояние возбуждения, когда на короткое время мембрана становится проницаемой и для других ионов, в частности, для натрия и хлора. В результате притока внутрь клетки положительных ионов величина мембранного потенциала падает, а затем, после восстановления избирательных свойств мембраны, снова входит в норму. Так впервые была объяснена генерация спайка.
. Далее, Бернштейн предположил, что в жизни нейрона иногда наступает особый момент -состояние возбуждения, когда на короткое время мембрана становится проницаемой и для других ионов, в частности, для натрия и хлора. В результате притока внутрь клетки положительных ионов величина мембранного потенциала падает, а затем, после восстановления избирательных свойств мембраны, снова входит в норму. Так впервые была объяснена генерация спайка.
Натриево - калиевый цикл
Гипотеза Бернштейна хорошо объясняла многие известные физиологам того времени факты. В частности, зависимость потенциала покоя от концентрации калия в межклеточной среде, а также от температуры. Однако в то время гипотезу нельзя было подвергнуть экспериментальной проверке. В распоряжении исследователей не было методики непосредственного изменения мембранного потенциала и концентраций ионов. Гипотеза Бернштейна вызывала массу споров.
Ситуация изменилась после сообщения английского зоолога Дж. Юнга, сделанного в 1936 г. Он обнаружил, что длинные тяжи у кальмаров и каракатиц являются не кровеносными сосудами, как это считалось ранее, а необычайно толстыми аксонами (отростками нервных клеток). Они получили название гигантских аксонов и стали естественным объектом для изучения мембран. Диаметр аксонов достигает 1 мм и дает возможность вводить в них электроды и капилляры. В результате можно измерить как мембранный потенциал, так и концентрацию внутриклеточного раствора.
Выяснилось, что мембранная теория Бернштейна в целом верна, хотя и нуждается во внесении ряда существенных дополнений и изменений. В частности верным оказалось утверждение, что потенциал покоя обусловлен в основном
электрохимическим потенциалом ионов ![]() . Наоборот, согласно теории Бернштейна в точке максимума потенциала действия (спайка) мембранный потенциал должен быть равен нулю (ток ионов прекращается, когда напряжение равно нулю). Однако, английские физиологи А. Ходжкин и А. Хаксли в 1939 г. показали, что на максимуме потенциала действия мембранный потенциал не уничтожается, а меняет знак. Внешняя поверхность мембраны заряжается положительно по отношению к внутренней. Явление получило название “овершут”
и не имело объяснений в рамках теории Бернштейна. Оно связано с неравномерным распределением ионов натрия. Оказалось, что концентрация ионов
. Наоборот, согласно теории Бернштейна в точке максимума потенциала действия (спайка) мембранный потенциал должен быть равен нулю (ток ионов прекращается, когда напряжение равно нулю). Однако, английские физиологи А. Ходжкин и А. Хаксли в 1939 г. показали, что на максимуме потенциала действия мембранный потенциал не уничтожается, а меняет знак. Внешняя поверхность мембраны заряжается положительно по отношению к внутренней. Явление получило название “овершут”
и не имело объяснений в рамках теории Бернштейна. Оно связано с неравномерным распределением ионов натрия. Оказалось, что концентрация ионов ![]() внутри клетки примерно в десять раз ниже чем в межклеточной жидкости. В начале спайка ионы натрия под действием электрического поля и концентрационного градиента устремляются из межклеточной среды в клетку (в клетке их концентрация ниже, а внутренняя поверхность мембраны заряжена отрицательно по отношению к внешней). В этом гипотеза Бернштейна оказалась верной. После смены знака мембранного потенциала ионы натрия продолжают перемещаться в клетку только под действием концентрационного градиента. Мембранный потенциал на максимуме овершута приближается к равновесному электрохимическому потенциалу для ионов
внутри клетки примерно в десять раз ниже чем в межклеточной жидкости. В начале спайка ионы натрия под действием электрического поля и концентрационного градиента устремляются из межклеточной среды в клетку (в клетке их концентрация ниже, а внутренняя поверхность мембраны заряжена отрицательно по отношению к внешней). В этом гипотеза Бернштейна оказалась верной. После смены знака мембранного потенциала ионы натрия продолжают перемещаться в клетку только под действием концентрационного градиента. Мембранный потенциал на максимуме овершута приближается к равновесному электрохимическому потенциалу для ионов ![]() . В дальнейшем проницаемость мембраны для ионов натрия падает и активируется проводимость для ионов калия, которые движутся из клетки, где их концентрация выше. Ионы калия уносят из клетки положительный заряд. Внутренняя поверхность мембраны заряжается отрицательно. Этот натриево-калиевый цикл ниже будет рассмотрен подробнее.
. В дальнейшем проницаемость мембраны для ионов натрия падает и активируется проводимость для ионов калия, которые движутся из клетки, где их концентрация выше. Ионы калия уносят из клетки положительный заряд. Внутренняя поверхность мембраны заряжается отрицательно. Этот натриево-калиевый цикл ниже будет рассмотрен подробнее.
В течение цикла ионы натрия заходят в клетку, а ионы калия ее покидают. Такое замещение должно выравнивать концентрации ионов. Казалось бы, что после ряда циклов нервная клетка не должна генерировать спайков (концентрации ионов внутри и вне клетки уравняются). В действительности это не так. Неравномерное распределение ионов внутри и вне клетки поддерживается активным транспортом ионов. Механизмы, осуществляющие транспорт ионов через мембрану в сторону повышенной концентрации, называются мембранными насосами (помпами). Они потребляют энергию, которая освобождается в результате окисления аминокислот.
Наиболее изучен натриевый насос. Он выводит ионы ![]() из клетки и одновременно транспортирует ионы
из клетки и одновременно транспортирует ионы ![]() внутрь клетки. Этот процесс включает в себя обязательный обмен двух ионов
внутрь клетки. Этот процесс включает в себя обязательный обмен двух ионов ![]() во внешней среде на три иона
во внешней среде на три иона ![]() в клетке (если удалить из среды ионы
в клетке (если удалить из среды ионы ![]() , то ионы
, то ионы ![]() из клетки выводиться уже не будут). Если мембранный потенциал отрицателен (внутренняя поверхность мембраны заряжена отрицательно), то натриевый насос выполняет электрическую работу, выводя из клетки суммарно один положительный заряд. Натриевый насос действует избирательно. Например, он не способен переносить ионы лития. Молекулярный механизм активного транспорта в настоящее время до конца не ясен.
из клетки выводиться уже не будут). Если мембранный потенциал отрицателен (внутренняя поверхность мембраны заряжена отрицательно), то натриевый насос выполняет электрическую работу, выводя из клетки суммарно один положительный заряд. Натриевый насос действует избирательно. Например, он не способен переносить ионы лития. Молекулярный механизм активного транспорта в настоящее время до конца не ясен.
Натриевый насос - один из главных механизмов, благодаря которому внутри клетки поддерживается избыточная по сравнению с внешней средой концентрация ионов ![]() и недостаточная, примерно в десять раз меньшая, чем во внеклеточной жидкости, концентрация ионов
и недостаточная, примерно в десять раз меньшая, чем во внеклеточной жидкости, концентрация ионов ![]() .
.
Для исследования процесса генерации спайка А.Ходжкиным и А.Хаксли был разработан оригинальный прием, который называется методом фиксации потенциала. Внутриклеточно вводятся электроды и используется система с обратной связью, позволяющая поддерживать мембранный потенциал на любом заданном уровне. Тем самым, можно имитировать различные фазы потенциала действия, как бы “замораживая” его. Система позволяет измерять ионные токи. Изменяя ионный состав среды (так же через микрокапилляры меняют состав внутриклеточной жидкости) можно идентифицировать ионы, которые обуславливают ток. Исследования обнаружили, что даже пассивный транспорт ионов (в соответствии с градиентом концентраций и направлением электрического поля) подчиняется специфическим закономерностям. Для их объяснения А.Ходжкиным и А.Хаксли была высказана гипотеза о наличии в мембране специальных ионных каналов, служащих для их транспортировки.
При положительном отклонении мембранного потенциала от потенциала в состоянии покоя (значение потенциала уменьшается по абсолютной величине) активируются, т.е. открываются каналы для ![]() , пропускающие эти ионы внутрь клетки. Ионы
, пропускающие эти ионы внутрь клетки. Ионы ![]() движутся в соответствии с направлением электрического поля, причем в область с меньшей концентрацией. Каналы играют роль селективных (избирательных) фильтров, пропускающих ионы
движутся в соответствии с направлением электрического поля, причем в область с меньшей концентрацией. Каналы играют роль селективных (избирательных) фильтров, пропускающих ионы ![]() и непроницаемых для других ионов. Механизмы, приводящие к открытию и закрытию каналов, называются воротами.
При деполяризации (уменьшении абсолютной величины мембранного потенциала) открываются активационные так называемые
и непроницаемых для других ионов. Механизмы, приводящие к открытию и закрытию каналов, называются воротами.
При деполяризации (уменьшении абсолютной величины мембранного потенциала) открываются активационные так называемые ![]() -ворота для натриевых каналов. Чем сильнее деполяризована мембрана, тем больше
-ворота для натриевых каналов. Чем сильнее деполяризована мембрана, тем больше ![]() -ворот открыто, что в свою очередь увеличивает деполяризацию. Процесс открытия ворот протекает лавинообразно.
-ворот открыто, что в свою очередь увеличивает деполяризацию. Процесс открытия ворот протекает лавинообразно.
При относительно длительной деполяризации натриевые каналы инактивируются. Срабатывают так называемые ![]() -ворота, которые блокируют каналы через время порядка 1 мсек после открытия
-ворота, которые блокируют каналы через время порядка 1 мсек после открытия ![]() -ворот. В 1980 г. Ф. Сигворс и Э. Неер зарегистрировали токи через единичные натриевые каналы. Оказалось, что они подчиняются закону “все или ничего”, Открываются практически мгновенно, а через время около 1 мсек. так же практически мгновенно закрываются. Ионные каналы представляют собой молекулы белка, вкрапленные в липидный слой мембраны.
-ворот. В 1980 г. Ф. Сигворс и Э. Неер зарегистрировали токи через единичные натриевые каналы. Оказалось, что они подчиняются закону “все или ничего”, Открываются практически мгновенно, а через время около 1 мсек. так же практически мгновенно закрываются. Ионные каналы представляют собой молекулы белка, вкрапленные в липидный слой мембраны.
В районы максимума пика потенциала действия большинство натриевых каналов инактивируется, т.е. закрываются ![]() -ворота. Встает вопрос: каким образом мембранный потенциал возвращается к исходному уровню -состоянию покоя? Экспериментально А. Ходжкин и А. Хаксли обнаружили, что с некоторым запаздыванием по отношению к началу натриевого тока возникает калиевый ток. Ионы калия начинают покидать клетку, поскольку активируются калиевые каналы. Гипотеза связывает это с открытием так называемых
-ворота. Встает вопрос: каким образом мембранный потенциал возвращается к исходному уровню -состоянию покоя? Экспериментально А. Ходжкин и А. Хаксли обнаружили, что с некоторым запаздыванием по отношению к началу натриевого тока возникает калиевый ток. Ионы калия начинают покидать клетку, поскольку активируются калиевые каналы. Гипотеза связывает это с открытием так называемых ![]() -ворот. Ионы калия движутся в область с меньшей концентрацией (пассивный транспорт). При этом в районе овершута (мембранный потенциал положителен) движение осуществляется в соответствии с направлением электрического поля. Калиевый ток уносит из клетки (точнее с внутренней поверхности мембраны) отрицательный заряд. Мембрана снова поляризуется.
-ворот. Ионы калия движутся в область с меньшей концентрацией (пассивный транспорт). При этом в районе овершута (мембранный потенциал положителен) движение осуществляется в соответствии с направлением электрического поля. Калиевый ток уносит из клетки (точнее с внутренней поверхности мембраны) отрицательный заряд. Мембрана снова поляризуется.
Считается, что в отличие от натриевых каналов, для калиевых каналов нет инактивационных ворот. Активационные ![]() -ворота сами закрываются, когда мембрана возвращается к уровню потенциала покоя (реполяризуется).
-ворота сами закрываются, когда мембрана возвращается к уровню потенциала покоя (реполяризуется).
Важна отметить, что количество ионов, которое проходит через мембрану при одиночном потенциале действия, практически не вызывает изменения внутриклеточных концентраций ионов. Исключение могут составлять лишь мельчайшие нервные клетки. По некоторым оценкам при одном потенциале действия содержание ![]() внутри гигантского аксона кальмара изменяется всего лишь на
внутри гигантского аксона кальмара изменяется всего лишь на ![]() от исходного. Если блокировать активные натриевые насосы каким-нибудь ядом, то аксон все равно сможет сгенерировать несколько тысяч импульсов. Непосредственно активный транспорт не участвует в генерации спайков. Однако, он поддерживает на должном уровне концентрационные градиенты, наличие которых приводит к возникновению потенциалов действия.
от исходного. Если блокировать активные натриевые насосы каким-нибудь ядом, то аксон все равно сможет сгенерировать несколько тысяч импульсов. Непосредственно активный транспорт не участвует в генерации спайков. Однако, он поддерживает на должном уровне концентрационные градиенты, наличие которых приводит к возникновению потенциалов действия.
После описания механизма генерации спайка, обратимся к системе уравнений, описывающей это процесс. Она предложена А. Ходжкиным и А. Хаксли.
Аксон Ходжкина - Хаксли
Модель базируется на электрохимическом уравнении (3). Мембранный потенциал будем отсчитывать от его уровня ![]() в состоянии покоя, т.е. положим
в состоянии покоя, т.е. положим ![]() , или же
, или же ![]() . Аналогично введем обозначения для отсчитываемых от
. Аналогично введем обозначения для отсчитываемых от ![]() равновесных электрохимических потенциалов
равновесных электрохимических потенциалов ![]() и
и ![]() . Ясно, что сохраняются разности:
. Ясно, что сохраняются разности: ![]() и
и ![]() . В создании мембранного потенциала кроме ионов
. В создании мембранного потенциала кроме ионов ![]()
29-04-2015, 01:59