Федеральное агентство по образованию
Государственное образовательное образовательное учреждение
Высшего профессионального образования
«Карельский государственный педагогический университет»
КОНТРОЛЬНАЯ РАБОТА
Физиология сенсорных систем.
Физиология Вкуса.
Выполнила: Войтенко Н.Д.
Студентка 2 курса ОЗО группа 823-3
Преподаватель: Зорова О.В.
Петрозаводск
2009 г.
Содержание
1. Физиология сенсорных систем
1.1. Периферический (рецепторный) отдел анализаторов……………………………….3
1.2. Проводникового отдела анализаторов………………………………………………...7
1.3. Центральный или корковый отдел анализатора……………………………………...8
2. Физиология вкуса
2.1. Морфология органов вкуса; субъективная физиология вкуса. Ориентация и строение вкусовых почек………………………………………………………………9
2.2. Центральные связи…………………………………………………………………....11
2.3. Основные вкусовые ощущения………………………………………………………11
2.4. Интенсивность ощущений……………………………………………………………12
2.5. Объективная физиология вкуса………………………………………………………13
2.6. Первичный процесс…………………………………………………………………...13
2.7. Роль вкусовой чувствительности…………………………………………………….14
Литература………………………………………………………………………………………14
1. Физиология сенсорных систем
Человек и животное непрерывно получают информацию о бесконечном многообразии изменений, которые происходят во внешней и внутренней среде. Это осуществляется благодаря наличию у организма специализированных структур, которые получили название анализаторы (сенсорные системы).
Под анализаторами понимают совокупность образований, обеспечивающих восприятие энергии раздражителя, трансформацию ее в специфические процессы возбуждения, проведение этого возбуждения в структуры ЦНС и к клеткам коры, анализ и синтез специфическими зонами коры этого возбуждения с последующим формированием ощущения.
Понятие об анализаторах введено в физиологию И. П. Павловым в связи с учением о высшей нервной деятельности. Каждый анализатор состоит из трех отделов:
Периферический или рецепторный отдел, который осуществляет восприятие энергии раздражителя и трансформацию ее в специфический процесс возбуждения.
Проводниковый отдел, представленный афферентными нервами и подкорковыми центрами, он осуществляет передачу возникшего возбуждения в кору головного мозга.
Центральный или корковый отдел анализатора, представленный соответствующими зонами коры головного мозга, где осуществляется высший анализ и синтез возбуждений и формирование соответствующего ощущения.
Роль анализаторов при формировании приспособительных реакций чрезвычайно велика и многообразна. Согласно концепции функциональной системы П. К. Анохина формирование любой приспособительной реакции осуществляется в несколько этапов. Анализаторы принимают непосредственное участие в формировании всех этапов функциональной системы. Они являются поставщиками афферентных посылок определенной модальности и различного функционального назначения, причем, одна и та же афферентация может быть обстановочной, пусковой, обратной и ориентировочной в зависимости от этапа формирования приспособительной деятельности.
1.1 Периферический (рецепторный) отдел анализаторов
Рецепторы играют ведущую роль в получении организмом информации о состоянии внешней и внутренней среды. Благодаря большому многообразию рецепторов человек способен воспринимать стимулы разных модальностей.
Рецепторы представляют собой конечные специализированные образования, которые предназначены для восприятия энергии раздражителя и трансформации ее в специфическую активность нервной клетки. У большинства рецепторных аппаратов основной структурной единицей является клетка, снабженная подвижными волосками или ресничками, которые представляют собой как бы периферические подвижные антенны. В составе волосков выделяют 9 пар периферических фибрилл, которые сокращаются под действием атф, благодаря чему осуществляется непрерывные поиски адекватного стимула и обеспечиваются условия взаимодействия с ним. Центральные 2 фибриллы выполняют опорную функцию.
Общий механизм рецепции слагается из механохимических молекулярных процессов, обеспечивающих движение антенн, и общих биохимических циклов при взаимодействии специфического стимула с рецепторными мембранами антенн. Однако, у некоторых рецепторов во взаимодействии со стимулом принимает участие вся клетка (например, хеморецепторные клетки, чувствительные к напряжению кислорода в крови), у других восприятие осуществляется микроворсинками (вкусовые луковицы). У большинства рецепторов кожи, внутренних органов и мышц участки преобразования стимула находятся в окончаниях нервных волокон.
Классификация рецепторов. В основу классификации рецепторов положено несколько критериев.
Психофизиологический характер ощущения: тепловые, холодовые, болевые и др.
Природа адекватного раздражителя: механо-, термо-, хемо-, фото-, баро-, осмбрецепторы и др.
Среда, в которой рецептор воспринимает раздражитель: экстеро-, интерорецепторы.
Отношение к одной или нескольким модальностям: моно- и полимодальные (мономодальные преобразуют в нервный импульс только один вид раздражителя - световой, температурный и т. д., полимодальные могут несколько раздражителей преобразовать в нервный импульс - механический и температурный, механический и химический и т. д.).
Способность воспринимать раздражитель, находящийся на расстоянии от рецептора или при непосредственном контакте с ним: контактные и дистантные.
Уровень чувствительности (порог раздражения): низкопороговые (механорецепторы) и высокопороговые (ноцицепторы).
Скорость адаптации: быстроадаптирующиеся, (тактильные), медленноадаптирующиеся (болевые) и неадаптирующиеся (вестибулярные рецепторы и проприорецепторы).
Отношение к различным моментам действия раздражителя: при включении раздражителя, при его выключении, на протяжении всего времени действия раздражителя.
Морфофункциональная организация и механизм возникновения возбуждения: первичночувствующие и вторичночувствующие.
В первичночувствующих рецепторах стимул действует на воспринимающий субстрат, заложенный в самом сенсорном нейроне, который при этом возбуждается непосредственно (первично) раздражителем. К первичночувствующим рецепторам относятся: обонятельные, тактильные рецепторы и мышечные веретена.
К вторичночувствующим относятся те рецепторы, у которых между действующим стимулом и сенсорным нейроном располагаются дополнительные рецептирующие клетки, при этом сенсорный нейрон возбуждается не непосредственно стимулом, а опосредовано (вторично) - потенциалом рецептирующей клетки. К вторичночувствующим рецепторам относятся: рецепторы слуха, зрения, вкуса, вестибулярные рецепторы.
Механизм возникновения возбуждения у этих рецепторов различен. В первичночувствующем рецепторе транформация энергии раздражителя и возникновение импульсной активности идет в самом сенсорном нейроне. У вторичночувствующих рецепторов между сенсорным нейроном и стимулом расположена рецептирующая клетка, в которой под влиянием раздражителя идут процессы трансформации энергии раздражителя в процесс возбуждения. Но в этой клетке не возникает импульсной активности. Рецепторные клетки синапсами соединены с сенсорными нейронами. Под влиянием потенциала рецептирирующей клетки выделяется медиатор, который возбуждает нервное окончание сенсорного нейрона и вызывает в нем появление локального ответа - постсинаптического потенциала. Он оказывает деполяризующее действие на отходящее нервное волокно, в котором возникает импульсная активность.
Свойства периферического (рецепторного) отдела анализаторов. В деятельности каждого анализатора и его отделов независимо от характеристики раздражителей различают ряд общих свойств. Для периферического отдела анализаторов характерны следующие свойства.
1. Специфичность - способность воспринимать определенный, т. е. адекватный данному рецептору, раздражитель. Эта способность рецепторов сформировалась в процессе эволюции.
2. Высокая чувствительность - способность реагировать на очень малые по интенсивности параметры адекватного раздражителя. Например, для возбуждения фоторецепторов сетчатки глаза достаточно нескольких, а иногда и одного, квантов света. Обонятельные рецепторы информируют организм о появлении в атмосфере единичных молекул пахучих веществ.
3. Способность к ритмической генерации импульсов возбуждения в ответ на однократное действие раздражителя.
4. Способность к адаптации - т. е. способность приспосабливаться ("привыкать") к постоянно действующему стимулу. Адаптация может выражаться в снижении активности рецептора и частоты генерации импульсов возбуждения, вплоть до полного его прекращения. В зависимости от скорости адаптации различают:
быстроадаптирующиеся (тактильные);
медленноадаптирующиеся (терморецепторы);
неадаптирующиеся (вестибулярные и проприорецепторы). Выделяют несколько видов адаптации:
изменение возбудимости рецептора в сторону снижения - десенсибилизация;
изменение возбудимости в сторону повышения - сенсибилизация.
Адаптация проявляется в снижении абсолютной чувствительности рецептора и в повышении дифференциальной чувствительности к стимулам, близким по силе к адаптируемому. Сенсибилизация проявляется в стойком повышении возбудимости, которое вызывается многократными действиями пороговых раздражителей, наносимых один за другим.
5. Функциональная мобильность. Анализаторные системы способны изменять свою деятельность путем изменения количества функционирующих рецепторов в зависимости от условий окружающей среды и функционального состояния организма. Например, количество функционирующих вкусовых рецепторов больше в состоянии голода, а после приема пищи их количество уменьшается. При снижении температуры окружающей среды количество холодовых рецепторов кожных покровов увеличивается.
6. Низкая способность к аккомодации.
7. Специализация рецепторов к определенным параметрам адекватного раздражителя. Рецепторы, входящие в состав периферического отдела анализатора, неоднородны по отношению к различным моментам действия раздражителя. Имеются рецепторы, которые возбуждаются только в момент включения раздражителя, другие- только в момент выключения раздражителя, а третьи реагируют в течение всего времени действия раздражителя. Кроме того, имеются рецепторы, реагирующие на изменение интенсивности раздражителя или на его перемещение и т. д.
8. Способность к элементарному первичному анализу. Благодаря связи между отдельными рецепторами периферического отдела, отражающими отдельные параметры раздражителя, осуществляется элементарный первичный анализ последнего. Деятельность рецепторов осуществляется не изолированно, а во взаимодействии, в связи с чем уже на рецепторном уровне осуществляется анализ раздражителя по разным его характеристикам (параметрам).
9. Кодирование информации. Информация о действии химических, механических раздражителей, имеющих разнообразную природу, преобразуется рецепторами в универсальные для мозга сигналы - нервные импульсы. Таким образом рецепторы кодируют информацию о среде, т. е. преобразуя сигналы, непонятные мозгу, в сигналы, понятные ему.
Кодирование качества. Различение действующих на организм внешних раздражителей по их физической и химической природе происходит уже при первой встрече с ними соответствующих рецепторов. Это различение достигается избирательной чувствительностью рецепторов к определенному виду энергии и очень низкими порогами возбуждения.
Кодирование интенсивности. Так как частота афферентной импульсации зависит от амплитуды рецепторного потенциала, которая в свою очередь пропорциональна интенсивности раздражения, то кодирование интенсивности стимула осуществляется посредством изменения частоты следования нервных импульсов от рецепторов в нервные центры. Увеличение интенсивности раздражителя кодируется увеличением частоты импульсной активности.
Между интенсивностью стимула и частотой потенциалов действия существует логарифмическая зависимость - ощущение увеличивается пропорционально логарифму интенсивности раздражения. Эта зависимость получила название закона Вебера-Фехнера, описавших ее.
Пространственное кодирование. В некоторых сенсорных системах естественная стимуляция рецепторов характеризуется тем или иным распределением локальных стимулов. Способность определять место или конфигурацию стимулов называется пространственным различением. В зрительной и слуховой системах выделены афферентные каналы, пространственно разнесенные в центральных структурах и связанные с обработкой информации о локализации источника раздражения, его перемещении, хроматических и частотных качествах сигнала.
Временное кодирование. Способность оценки времени неотделима от других аспектов кодирования. Частота нервных разрядов - это универсальная переменная величина, которая изменяется во времени. Кодирование информации осуществляется группой равномерно следующих импульсов. В качестве сигнальных признаков используются такие временные параметры выходных сигналов, как частота импульсации или продолжительность межимпульсных интервалов. Для временного различия двух раздражителей необходимо, чтобы нервные процессы, вызванные этими раздражителями, не сливались во времени.
Таким образом, уже на уровне рецепторов осуществляется первичное кодирование качества стимулов и их количественных характеристик - переход из присущей им формы физической и химической энергии в форму нервных импульсов. Преобразованная информация поступает на следующий уровень сенсорной системы, где подвергается дальнейшим преобразованиям, приводящим к изменению кода. Ни на одном уровне сенсорной системы не происходит восстановления стимула в его первоначальной форме, т. е. декодирование. Это основное отличие физиологического кодирования от большинства технических систем связи, где сообщение, как правило, восстанавливается в первоначальном, декодированном виде.
1.2 Проводниковый отдел анализаторов
Этот отдел анализаторов представлен афферентными путями и подкорковыми центрами. Основными функциями проводникового отдела являются: анализ и передача информации, осуществление рефлексов и межанализаторного взаимодействия. Эти функции обеспечиваются свойствами проводникового отдела анализаторов, которые выражаются в следующем.
1. От каждого специализированного образования (рецептора), идет строго локализованный специфический сенсорный путь. Эти пути как правило, передают сигналы от рецепторов одного типа.
2. От каждого специфического сенсорного пути отходят коллатерали к ретикулярной формации, в результате чего она является структурой конвергенции различных специфических путей и формирования мультимодальных или неспецифических путей, кроме того, ретикулярная формация является местом межанализаторного взаимодействия.
3. Имеет место многоканальность проведения возбуждения от рецепторов к коре (специфические и неспецифичекие пути), что обеспечивает надежность передачи информации.
4. При передаче возбуждения происходит многократное переключение возбуждения на различных уровнях ЦНС. Выделяют три основных переключающих уровня:
спинальный или стволовой (продолговатый мозг);
зрительный бугор;
соответствующая проекционная зона коры головного мозга.
Вместе с тем, в пределах сенсорных путей существуют афферентные каналы срочной передачи информации (без переключении) в высшие мозговые центры. Полагают, что по этим каналам осуществляется преднадстройка высших мозговых центров к восприятию последующей информации. Наличие таких путей является признаком совершенствования конструкции мозга и повышения надежности сенсорных систем.
5. Кроме специфических и неспецифических путей существуют так называемые ассоциативные таламо-кортикальные пути, связанные с ассоциативными областями коры больших полушарий. Показано, что с деятельностью таламо-кортикальных ассоциативных систем связана межсенсорная оценка биологической значимости стимула и др. Таким образом, сенсорная функция осуществляется на основе взаимосвязанной деятельности специфических, неспецифических и ассоциативных образований мозга, которые и обеспечивают формирование адекватного адаптивного поведения организма.
1.3 Корковый отдел анализаторов
1. Каждая сенсорная система (каждый анализатор) имеет проекцию в кору больших полушарий. Корковый отдел анализаторов имеет центральную часть и окружающую ее ассоциативную зону (по представлению И. П. Павлова - "ядро" и рассеянные элементы). Центральная часть коркового отдела анализатора состоит из высокодифференцированных в функциональном отношении нейронов, которые осуществляют высший анализ и синтез информации, поступающей к ним. Ассоциативные корковые зоны представлены менее дифференцированными нейронами, способных к выполнению простейших функций. Синтез и анализ афферентных импульсов этими клетками осуществляется в элементарной, примитивной форме.
2. Одной из общих черт организации сенсорных систем является принцип двойственной проекции их в кору больших полушарий. Этот принцип тесно связан с многоканальностью проводящих путей и выражается в осуществлении двух различных типов корковых проекций, которые можно разделить на первичные и вторичные проекции. Первичные и вторичные проекционные зоны окружены ассоциативными корковыми зонами той же сенсорной системы. Примером двойственной проекции в коре головного мозга может служить представительство вкусового анализатора. Его первичная корковая проекция представлена, по-видимому, орбитальной областью коры, так как именно здесь при раздражении рецепторов языка вызванные ответы возникают с самым коротким латентным периодом и имеют самую высокую амплитуду. Вторичной проекционной областью коры вкусового анализатора является соматосенсорная область. Здесь вызванные ответы возникают значительно позже, чем в орбитальной области, и амплитуда их меньше.
3. Взаимодействие анализаторов на корковом уровне осуществляется за счет ассоциативных корковых зон и за счет наличия полимодальных нейронов. Деятельность одних анализаторов находится в зависимости от деятельности других, причем, может наблюдаться как усиление деятельности анализатора, так и ее ослабление.
2. ФИЗИОЛОГИЯ ВКУСА
2.1.Морфология органов вкуса; субъективная физиология вкуса. Ориентация и строение вкусовых почек.
Язык у человека покрыт слизистой оболочкой, складки которой во многих местах образуют маленькие выпуклости в форме колышков, называемые сосочками. На рис.1 показано распределение трех типов сосочков-желобоватых, листовидных и грибовидных-по поверхности языка.
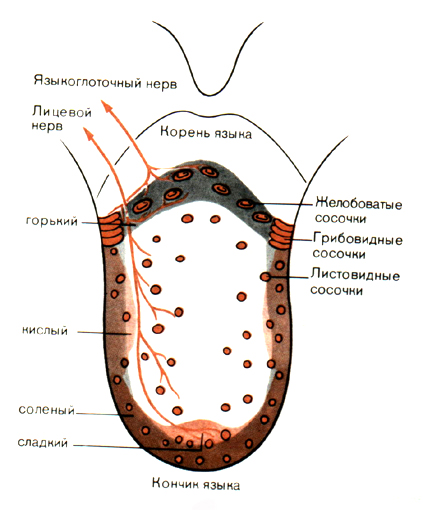
Рис. 1. Схема распределения вкусовых сосочков, их иннервации и зон максимальной чувствительности к разным вкусовым качествам на языке человека. [2]
Эти три типа распределены по-разному. Только грибовидные сосочки рассеяны по всей поверхности. Желобоватые сосочки, которых у человека всего 7-12, сверху имеют вид круглых образований 1-3 мм в диаметре; они находятся в ограниченной зоне поперек спинки языка у его корня. Третий тип, листовидные сосочки, образуют тесно расположенные складки вдоль задних краев языка. Они хорошо развиты у детей, но гораздо менее выражены и менее многочисленны у взрослых.
Нитевидные сосочки, занимающие остальную поверхность языка, не показаны на рис. 1, потому что в них нет вкусовых почек. Название "почка" говорит о форме этих органов (рис. 2). Положение их на сосочках варьирует; в случае желобоватых и листовидных сосочков много вкусовых почек заложено в боковых стенках, а на верхушке их нет. В грибовидных сосочках вкусовые почки ограничены поверхностью "шляпки гриба", которая может достигать 1 мм в диаметре.
Отдельная вкусовая почка имеет около 70 мкм в высоту и около 40 мкм в диаметре. Всего у человека около 2000 вкусовых почек, из них около половины-на желобоватых сосочках. Каждая вкусовая почка содержит 40-60 отдельных клеток.
В соединительную ткань под желобоватыми и листовидными сосочками погружены серозные железы, протоки которых открываются в углубления у основания сосочка, их секрет служит для смывания частиц пищи и микроорганизмов. Кроме того, он понижает концентрацию стимулирующего вещества вблизи вкусовых почек.
Внутри вкусовых почек различают три типа клеток: сенсорные, опорные и базальные (рис. 2). Растворимые в воде вещества, попадающие на поверхность языка, диффундируют через пору в наполненное жидкостью пространство над вкусовой почкой; здесь они соприкасаются с мембранами микровиллей, которые образуют наружные концы сенсорных клеток. Вкусовые рецепторы представляют собой вторичные сенсорные клетки без аксонов, которые проводят импульсы в центральном направлении. Их ответы передаются афферентными волокнами, которые образуют синапсы близ оснований сенсорных клеток. На рис. 2 показаны только два волокна, но в действительности в каждую вкусовую почку входят и разветвляются в ней около 50 волокон.
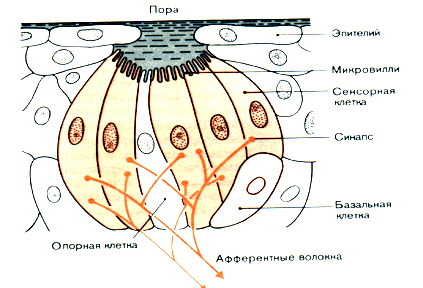
Рис. 2. Строение и иннервация вкусовой почки.
Продолжительность жизни сенсорных клеток во вкусовых почках невелика; происходит их непрерывная смена. В среднем одна сенсорная клетка замещается новой уже через 10 дней. За сменой клеток можно проследить, помечая их ядра 3Н-тимидином и определяя число меченых ядер, сохранившихся через некоторое время. Утраченные сенсорные клетки замещаются новыми, которые образуются из базальных клеток . При этой смене должны прерваться синапсы между афферентными волокнами и старыми клетками и возникнуть новые синапсы. В связи с такой перестройкой возникает много интересных вопросов, особенно если учесть тот факт, что сенсорные клетки различаются по своей чувствительности к разным стимулам. Так, смена сенсорных клеток может привести к изменению "вкусового профиля"-характерной формы ответов в афферентных волокнах, о чем пойдет речь в следующем разделе.
2.2 Центральные связи.
Афферентные волокна, проводящие ответы от скоплений вкусовых луковиц, распределяются по двум черепномоз-говым нервам -лицевому (VII) и языкоглоточному (IX). Такое деление обычно соответствует областям языка, которые снабжаются этими волокнами. Так, волокна от желобоватых и листовидных сосочков идут преимущественно в составе языкоглоточного нерва, а волокна от грибовидных сосочков в передней части языка входят в барабанную струну (chorda tympani), ветвь лицевого нерва. У детей имеются добавочные вкусовые органы в эпителии мягкого нёба и задней стенки глотки до гортани; они иннервируются главным образом блуждающим нервом (X).
В головном мозгу вкусовые
9-09-2015, 19:07