Індикаторами забруднення води важкими металами виступають Potamogeton perfoliatus і Potamogeton obtusitolius (залізо), Gliceria maxima (плюмбум); азотистими сполуками - Lecuna gibba, Meniophilum verticilatum; хлором - Gliceria maxima; індикаторами підвищення мінералізації води є Myriophilum spicatum, Sagittaria sagittifolia, підвищений вміст кальцію у воді і донних відкладеннях - Nymphoides peltata, Potamogeton crispus. Ці та деякі інші види в умовах забруднення води характеризується високою концентрації хімічних сполук і елементів у воді вегетують більш тривалий час, ніж інші.
Індикаторами евтрофування водойм, що відбувається під впливом антропогенних факторів, є Butomus umbellatus, Typha lafisolia ділянок, що не зазнають антропогенного впливу, - Ceratophyllum submersum;водойм з відносно чистою водою - Gliceria plekata, Potamogeton alpinus.
Гігрофіти слугують індикаторами акумулятивно – ерозійних процесів, що проходять в прибережній смузі водойм (Cuperus clifsorneis, Sagittaria platyphilla, Vallisneria spiralis). Вони індукують ділянки майбутнього обміління (Batrachium, Scirpus littoralis) і формування мулових відкладень (Alisma gramineum , Alisma plantago - aguatica), а також початкової стадії заростання водойм (Hydrocharis morsus–ranae, Polygonum amfibium). Є індикаторами процесів заболочення, які особливо характерні для водойм антропогенного евтрофорування і зниження рівня води (Carex panikulata, Egilisetum fluviatile, Hippuris vulgaris) [11].
Під само очищенням розуміють весь комплекс взаємозв’язку фізичних хімічних та біологічних процесів, в результаті діяльності яких якість води повертається до першочасткового стану [10].
Велику роль в процесах самоочищення забруднених вод відіграють вищі водні рослини, які розвиваються у великих кількостях на озерах і водосховищах, мілких річках, ставках.
За даними Кабанова (1958, 1961) зарості рослин можуть слугувати бар’єром при попаданні в водойму розсіяних забруднюючих речовин, а також пригнічують сапрофітну мікрофлору. Наприклад, у елодеї ясно виражені антагоністичні властивості по відношенню до сапрофітних бактерій, її розвиток може також прискорити відмирання дизентерійної палички у воді (Доливо – Добровільський, 1959).
Водні рослини часто грають значно більшу роль, ніж фітопланктон, в процесах фотосинтетичної реарегенерації (Вінберг, 1955).
Часто завислі частинки створюють значну каламутність води, що різко погіршує її санітарний стан та заважають очистці на водоочисних спорудах. Рослинність є бар’єром для завислих мінеральних та органічних сполук. При фільтрації води через зарослі напівводяних та занурених рослин її прозорість збільшується [3].
В заростях занурених рослин значно збільшується кількість розчиненого кисню, значно знижується кількість розчиненого кисню, значно знижується кількість сольового аміаку і наростає азот нітритів.
Вищі водні рослини та зообентос сприяють розвитку фітофільної фауни, яка також приймає участь у самоочищенні води та даних відкладень від органічних речовин, продуктів їх розпаду та бактерій.
Макрофіти використовують також у біологічному закріпленні берегів. Укріплення берегів за допомогою водних макролітів визнано найбільш надійним простим і екологічно вигідним заходом. В заростях макрофітів більшості прісноводних водойм відбувається нерест риб.
Але при використанні вищої водної рослинності можливостей і негативний вплив - при її відмиранні та розкладанні, вона стає джерелом вторинного забруднення. Є можливість заболочення водойми внаслідок розростання вищої водної рослинності. Тому необхідно своєчасно видалити їх з водойм.
Розділ 2. Основні гідрохімічні показники, їх характеристика
2.1 Температура
Молекулярна структура природної води характеризується взаємодією її простих молекул, внаслідок чого вона може бути у рідкому, твердому і газоподібному стані.
Для функціонування водних екосистем температурозалежні зміни фізико-хімічних властивостей води мають виняткове значення. Так, лід, який покриває водойми взимку, ізолює більш глибокі шари води від промерзання.
При різкому зниженні температури повітря замерзає лише поверхневий шар водойм, нижче якого температура води утримується на рівні плюсових значень. У той же час у неглибоких водоймах при сильних морозах може спостерігатись промерзання товщі води до самого дна. У таких випадках риби та інші водяні організми вмерзають у льодову товщу. Як правило, за таких умов відбувається їх масова загибель.
Але є випадки, коли такі риби, як карась, після нетривалого вмерзання і наступного розморожування залишаються живими. Це відбувається завдяки тому, що навколо тіла риб утворюється невеликий шар води з підвищеною концентрацією солей. За таких умов необхідна нижча температура для замерзання води. Що ж до забезпечення таких риб киснем, то в умовах різкого зниження метаболізму його мінімальні концентрації у воді дають змогу рибам перебувати певний час у стані анабіозу. Виживають також деякі безхребетні тварини планктону і бентосу, що утворюють біоценоз льоду — пагон.
При весняному підвищенні температури і таненні льоду після досягнення температури 4 °С вода з верхніх шарів як більш важка опускається на дно, а придонні шари піднімаються на поверхню. Внаслідок такої вертикальної циркуляції відбувається більш швидке прогрівання всієї товщі води, в тому числі і нижніх шарів, що позитивно впливає на перебіг біологічних процесів.
Для водних екосистем характерним є повільне охолодження і нагрівання води, чому сприяє надзвичайно висока теплоємність води. Це зумовлено витрачанням певної частини теплової енергії на розрив водневих зв'язків у асоційованих молекулах. При значному підвищенні температури повітря вода хоч і стає більш теплою, але її температура ніколи не зрівнюється з атмосферною внаслідок високої теплоти пароутворення. Так, питома теплота пароутворення (випаровування) при температурі 0 і 100 °С становить відповідно 2,5106 і 2,26106 Дж/кг. У спекотні дні зростає інтенсивність випаровування води, а відповідно і віддача тепла, що запобігає її перегріванню. І навпаки, при зниженні температури нижче 0°С і утворенні льоду вивільняється значна кількість тепла (373 Дж/кг), тому вода охолоджується повільно. Питома теплота плавлення льоду — 333 000 Дж/кг, його питома теплоємність при нормальному тиску і 0 °С дорівнює 2 120 Дж/кг*°С), а молекулярна теплопровідність при 0°С — 2,24 Дж/кг*°С. Для снігу цей показник становить 1,8, а для води — 0,6 Дж/кг*°С.
Завдяки високій теплоємності води діапазон коливань температури водного середовища, в якому мешкають гідробіонти, рідко коли перевищує 30—35 °С. У порівнянні з гідробіонтами наземні організми змушені пристосовуватись до значно більшого діапазону коливань температури навколишнього середовища.
Для визначення температури використовують водний ртутний термометр зі шкалою від 0 до 30-35°С з мітками 0,1-0,2°С. Коли для відбору проб користуються батометром, тоді температуру вімірюють термометром, який вмонтованa у батометр. Показання термометру знімають зразу після того, як витягнули батометр з води. Батометр витримують на заданій глибині не менше 3-5 хв. Температуру міряють з точнітю до 0,1 °С.
Якщо для проб використовують батометр без термометра, то температуру води на глибині 2-3 м міряють за допомогою термометра з чашечкою (рис. 1). Це водний термометр, вмонтований в металеву оправу з металевою чашкою знизу. У чашці знаходиться кінець термометру (резервуар зі ртуттю), туди крізь просвірлені зверху отвори втікає вода. Термометр з чашкою прикріплюють до розміченої палки і опускають на потрібну глибину, витримують там біля 5 хв. і швидко піднімають на поверхню. Можливо приблизно визначити температуру, коли опустити термометр у бутиль з водою, яка відбиралась з потрібної глибини. Для вимірювання температури самого верхнього шару води можна опустити термометр у ведро з водою, яка узята з поверхні.
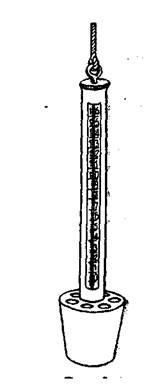
Рис. 1. Термометр з чашечкою.
2.2 Запах
Запах та смак води допомагає виявити сторонні забруднення. Навіть дуже незначні концентрації фенолу дають воді гострий запах карболової кислоти. Поганий запах води, який викликається сірководнем або особливими продуктами розкладу білкових речовин - меркаптанами (у яких кисень заміщенa сіркою) може вказувати на забруднення води фекальними скидами.
Визначення запаху досліджуваної води.
Бутиль, заповнену досліджуваною водою (на 3/4), щільно закривають, сильно перемішують, зразу відкривають і нюхають. Слабкий запах можна відчути при нагріванні. Досліджувану воду наливають у колбу, закривають Скляною пластинкою і нагрівають до 50°С (не більше). Потім піднімають пластинку та нюхають воду.
Для визначення смаку особливих вказівок не треба. Інтенсивність запаху та смаку можна характеризувати словами: без запаху та смаку, слабкий, помітний, сильний, дуже сильний. При цьому вказують, який саме запах має вода: невизначений, болотний, затхлий, гнилостний, сірководневий і т.п.
2.3 Прозорість води
Прозорість води змінюється в залежності від сезону, кількості завислих частинок, глибини водойм та багатьох інших причин. У нестратифікованих водоймах прозорість води знижується у придонному шарі внаслідок зростання каламутності, яка пов'язана з порушенням донних ґрунтів, у стратифікованих найбільша прозорість води спостерігається у гіполімніоні, а найменша — у зоні максимального розвитку фітопланктону. Знижується прозорість води в зоні термоклину за рахунок більш високої її щільності та затримки детриту.
Прозорість води змінюється в залежності від кількості зважених в ній часток мінерального або органічного походження. Улітку, в період «цвітіння» водоймища, прозорість може дуже сильно знизитись, дякуючи великій кількості водоростей, які знаходяться у товщі води. Під час зимівлі, коли риба знаходиться у малорухомому стані, взмучені речовини мінерального походження можуть значно погіршити умови дихання і викликати збуджений стан у риби. Сильно каламутять воду стічні води крахмало-паточних і низки інших заводів. У цьому випадку зниження прозорості служить одним з показників забруднення водоймища зовні.
У Дніпрі та його водосховищах найбільша прозорість води спостерігається взимку, а найменша — під час весняної повені. Так, у Київському водосховищі в зимовий період вона становить 1,5—2,1 м, а в Каховському — 2,4—3,7 м.
Сонячна радіація проникає у воду лише на певну глибину. Основна її частина поглинається верхніми шарами води. Як уже відзначалось, при проходженні через атмосферу до поверхні Землі досягає сонячна енергія, яка дорівнює 150 Вт/м2. При проникненні у воду значна частина цієї енергії поглинається уже у верхніх шарах води. Так, при прозорості 0,7 м і висоті Сонця 12° поглинається однометровим шаром води близько 92 % енергії, при прозорості 1,2 м і висоті Сонця 39° — до 76 %, а при прозорості 1,6 і висоті Сонця 58° — до 46 %.
Негативно впливають на проникнення сонячної радіації у товщу води зарості вищих водяних рослин. Так, у порівнянні з відкритими акваторіями на поверхню води з рогозом широколистим (50—60 % заростевого покриття) надходить 40 %, а з очеретом звичайним (90—95 % заростевого покриття) — тільки 25 % сонячної радіації. Ще менше надходить сонячної енергії в товщу води. Наприклад, на глибині 0,3 м у заростях лепешняку плаваючого із заростевим покриттям 90 % надходить 15 %, а при покритті 60 % — 80 % сонячної енергії. Поглинання у верхніх шарах води значної частини сонячної радіації різко обмежує поширення у товщі води фотосинтезуючих рослин. Вони можуть розвиватись на відносно невеликій глибині у континентальних водоймах, морях і океанах.
Верхній шар води, в якому є достатня кількість світла для синтезу рослинами органічної речовини з використанням сонячної енергії, називається фотичним, нижній шар, куди не надходить сонячна енергія, — афотичним. Зона проникнення світла, в якій інтенсивність фотосинтезу перевищує інтенсивність дихання рослин, має назву евфотичної зони. її нижня межа, де фотосинтез урівноважує інтенсивність дихання, називається компенсаційним горизонтом.
У воді найінтенсивніше поглинаються довгі хвилі сонячної радіації, які енергетично найбільш близькі до відповідних параметрів фотобіологічних процесів. Випромінювання, що проникає крізь товщу прісних і морських вод, зосереджене переважно в блакитній частині спектру і має довжину хвиль 475 – 480 нм. У процесі фотобіологічних реакцій енергія сонячної радіації поглинається дискретними частинками, які називаються фотонами, або квантами. Фотосинтез у бактерій протікає у спектральному діапазоні 400 - 900 нм, вищих зелених рослин — 400 - 700 нм, водоростей — 400 - 550 нм. Якщо хвилі коротші від 300 нм, порушується молекулярна структура білків і нуклеїнових кислот, і відповідно — нормальне функціонування живих систем. Ось чому несуть загрозу біосфері скорочення і розриви озонового шару, який затримує проникнення на Землю саме таких квантів сонячного випромінювання.
При прозорості води найінтенсивніше процес фотосинтезу протікає на глибинах відповідно до 1,5 і 6, 3 м, де величина фотосинтетичної активної радіації не падає нижче 0,17 - 0,22 Дж/см2, оскільки для максимальної інтенсивності фотосинтезу необхідно саме така величина сонячної радіації.
Сонячна радіація відіграє виключно важливу роль у функціонуванні водних екосистем. З нею пов'язана поведінка і розселення гідробіонтів у біотопах. Серед них є організми, які інтенсивно розвиваються у верхніх шарах води, куди надходить найбільше сонячної енергії. Це переважно автотрофні організми: водорості, фотосинтетичні бактерії, вищі водяні рослини. У процесі фотосинтезу вони запасають велику кількість енергії у вигляді первинно продукованої органічної речовини (первинної продукції), яку потім використовують організми інших трофічних рівнів.
Інші процеси, які протікають за участю сонячної енергії, не пов'язані безпосередньо з перетворенням сонячного світла в енергію хімічних сполук. Світло може виступати як інформативний фактор, що керує поведінкою водяних рослин і тварин. Наприклад, фотоперіодичні реакції рослин, які синхронізують етапи репродуктивного циклу рослин, здійснюються за допомогою пігменту фітохрому [25].
У водяних тварин репродуктивні цикли теж пов'язані з фотоперіодичними реакціями, опосередкованими через пігментні системи. Такі реакції водяних організмів, як фототаксис та фототропізм, залежать від освітлення водойм.
Прозорість визначається безпосередньо на водоймищі за допомогою гладкого металевого диску, обов'язково білого кольору. Диск для вимірювання прозорості має по краям отвори, за допомогою яких його прикріплюють до тросу, який розмічений кольоровими лоскутами через кожні 10 см. Диск опускають у воду до тих пір, поки він не зникне з поля зору та піднімають, поки він знову стане помітним. Середня величина зникнення диску при зануренні і глибини появи його при підніманні і буде умовною величиною прозорості води, яку виражають у сантиметрах.
Каламутність визначається на глаз і характеризується якісно ( незначна, значна, осад при відстоюванні і т. д.) [30].
2.4 Кольоровість води
Кольоровість прісних вод визначається в умовних одиницях - градусах платиново-кобальтової шкали. Кольоровість води, яка перевищує 30°, береться як висока.
Колір природних вод залежить від забарвлення розчинених у ній речовин, завислих частинок та мікроорганізмів, що населяють водну товщу. Він зумовлений поєднанням властивостей водного середовища, берегів водойми та метеорологічними факторами. На колір води впливають завислі речовини автохтонного (внутрішньоводоймного) та алохтонного (надходять ззовні) походження [27].
Власний колір води (блакитний) виявляється лише у воді деяких чистих гірських озер. Цей колір залежить від вибіркового поглинання променів сонячного спектру: найперше у воді затухають найдовші світові хвилі (червоної частини спектру), а останніми — короткі хвилі його синьої частини. Сонячне світло, проходячи через воду, втрачає червоні промені, поступово перетворюючись з білого в синє.
Природні води можуть мати зелене, жовте, буре і навіть чорне забарвлення, що зумовлено переважно наявністю гумінових речовин.
Крім того, колір води часто змінюється внаслідок масового розвитку певних планктонних організмів («цвітіння»). Це так зване вегетаційне забарвлення. Зокрема, при «цвітінні» синьо-зеленими водоростями вода набуває відповідного кольору, а при їх розкладі — навіть темно-синього внаслідок виділення пігментів фікоціану і фікобілінів. Зелені водорості забарвлюють воду в зелений колір, діатомові надають їй жовтуватого відтінку, дуналіела солоноводна (Dunaliella salina), червона евглена (Euglena rubra) і примнезій маленький (Primnesium parvum) — червоного кольору.
Кольоровість прісних вод визначається порівнянням досліджуваного зразку зі стандартними розчинами, виготовленими з хлорплатинату калію (К2РtСl6) та хлористого кобальту (СоСl2*6Н2О). Можна також приготувати стандарт з К2Сr2О7 та хлористого кобальту. Зважують на аналітичних вагах 0,0872 г К2Сr2О7 і 2 г хлористого кобальту, розчиняють реактиви у невеликій кількості дистильованої води в мірній літровій колбі, додають 1 мл концентрованої сірчаної кислоти (H2SO4) та доводять дистильованою водою до 1 л. Кольоровість цього розчину дорівнює 500°. Стандарти для порівняння готують розведенням вихідного розчину дистильованою водою, 1 л якої вміщує 1 мл концентрованої сірчаної кислоти.
У випадку неможливості приготування повної шкали стандартів, кольоровість визначають у парних колориметричних циліндрах однакового діаметру з нефарбованого скла, поділених на мілілітри (від 1 до 100) і забезпечених знизу скляними кранами. В один з таких циліндрів наливають (до мітки) досліджувану воду, у інший - стандартний розчин, кольоровість якого не дуже відрізняється від кольоровості досліджуваної води (такий стандарт отримують розчиненням у 5, 10, 20 або 50 разів вихідного розчину в 500°). Дивлячись зверху униз (крізь увесь шар рідини) на білому фоні, зливають інтенсивніше зафарбовану рідину до тих пір, поки колір рідини в обох циліндрах не буде однаковим. Висота столбу рідини обернено пропорційна інтенсивності її зафарбування. Тому кольоровість досліджуваної води розраховують за формулою:
Кольоровість висота столба стандарту (мл)*кольоровість стандарту (градуси) (градуси) висота столба досліджуваної води (мл)
2.5 Вміст хімічних речовин
Під густиною розуміють масу одиничного об'єму води в кілограмах на 1 м3 — (кг/м3). Вона залежить від температури, наявності розчинних солей, а також від атмосферного тиску та вищерозташованих мас води.
Найбільша густина хімічно чистої води, яка практично не має розчинених солей, при температурі 3,98°С дорівнює 1 г/см3. Із зниженням температури до 0°С, коли вода ще не перетворилась у лід, її щільність становить 998,87 кг/м3. При переході у стан льоду вона відразу зменшується до 916,7 кг/м3. Падає густина і при підвищенні температури води вище 4°С. При досягненні 30°С
29-04-2015, 00:48