Метаболизм, или обмен веществ, химические превращения, протекающие от момента поступления питательных веществ в живой организм до момента, когда конечные продукты этих превращений выделяются во внешнюю среду. К метаболизму относятся все реакции, в результате которых строятся структурные элементы клеток и тканей, и процессы, в которых из содержащихся в клетках веществ извлекается энергия. Иногда для удобства рассматривают по отдельности две стороны метаболизма – анаболизм и катаболизм, т.е. процессы созидания органических веществ и процессы их разрушения. Анаболические процессы обычно связаны с затратой энергии и приводят к образованию сложных молекул из более простых, катаболические же сопровождаются высвобождением энергии и заканчиваются образованием таких конечных продуктов (отходов) метаболизма, как мочевина, диоксид углерода, аммиак и вода.
Термин «обмен веществ» вошел в повседневную жизнь с тех пор, как врачи стали связывать избыточный или недостаточный вес, чрезмерную нервозность или, наоборот, вялость больного с повышенным или пониженным обменом. Для суждения об интенсивности метаболизма ставят тест на «основной обмен». Основной обмен – это показатель способности организма вырабатывать энергию. Тест проводят натощак в состоянии покоя; измеряют поглощение кислорода (О2) и выделение диоксида углерода (СО2). Сопоставляя эти величины, определяют, насколько полно организм использует («сжигает») питательные вещества. На интенсивность метаболизма влияют гормоны щитовидной железы, поэтому врачи при диагностике заболеваний, связанных с нарушениями обмена, в последнее время все чаще измеряют уровень этих гормонов в крови.
Методы исследования. При изучении метаболизма какого-нибудь одного из питательных веществ прослеживают все его превращения от той формы, в какой оно поступает в организм, до конечных продуктов, выводимых из организма. В таких исследованиях применяется крайне разнообразный набор биохимических методов.
Использование интактных животных или органов. Животному вводят изучаемое соединение, а затем в его моче и экскрементах определяют возможные продукты превращений (метаболиты) этого вещества. Более определенную информацию можно получить, исследуя метаболизм определенного органа, например печени или мозга. В этих случаях вещество вводят в соответствующий кровеносный сосуд, а метаболиты определяют в крови, оттекающей от данного органа.
Поскольку такого рода процедуры сопряжены с большими трудностями, часто для исследования используют тонкие срезы органов. Их инкубируют при комнатной температуре или при температуре тела в растворах с добавкой того вещества, метаболизм которого изучают. Клетки в таких препаратах не повреждены, и так как срезы очень тонкие, вещество легко проникает в клетки и легко выходит из них. Иногда затруднения возникают из-за слишком медленного прохождения вещества сквозь клеточные мембраны. В этих случаях ткани измельчают, чтобы разрушить мембраны, и с изучаемым веществом инкубируют клеточную кашицу. Именно в таких опытах было показано, что все живые клетки окисляют глюкозу до СО2 и воды и что только ткань печени способна синтезировать мочевину.
Использование клеток. Даже клетки представляют собой очень сложно организованные системы. В них имеется ядро, а в окружающей его цитоплазме находятся более мелкие тельца, т.н. органеллы, различных размеров и консистенции. С помощью соответствующей методики ткань можно «гомогенизировать», а затем подвергнуть дифференциальному центрифугированию (разделению) и получить препараты, содержащие только митохондрии, только микросомы или прозрачную жидкость – цитоплазму. Эти препараты можно по отдельности инкубировать с тем соединением, метаболизм которого изучается, и таким путем установить, какие именно субклеточные структуры участвуют в его последовательных превращениях. Известны случаи, когда начальная реакция протекает в цитоплазме, ее продукт подвергается превращению в микросомах, а продукт этого превращения вступает в новую реакцию уже в митохондриях. Инкубация изучаемого вещества с живыми клетками или с гомогенатом ткани обычно не выявляет отдельные этапы его метаболизма, и только последовательные эксперименты, в которых для инкубации используются те или иные субклеточные структуры, позволяют понять всю цепочку событий.
Использование радиоактивных изотопов. Для изучения метаболизма какого-либо вещества необходимы: 1) соответствующие аналитические методы для определения этого вещества и его метаболитов; и 2) методы, позволяющие отличать добавленное вещество от того же вещества, уже присутствующего в данном биологическом препарате. Эти требования служили главным препятствием при изучении метаболизма до тех пор, пока не были открыты радиоактивные изотопы элементов и в первую очередь радиоактивный углерод 14C. С появлением соединений, «меченных» 14C, а также приборов для измерения слабой радиоактивности эти трудности были преодолены. Если к биологическому препарату, например к суспензии митохондрий, добавляют меченную 14C жирную кислоту, то никаких специальных анализов для определения продуктов ее превращений не требуется; чтобы оценить скорость ее использования, достаточно просто измерять радиоактивность последовательно получаемых митохондриальных фракций. Эта же методика позволяет легко отличать молекулы радиоактивной жирной кислоты, введенной экспериментатором, от молекул жирной кислоты, уже присутствовавших в митохондриях к началу эксперимента.
Хроматография и электрофорез. В дополнение к вышеупомянутым требованиям биохимику необходимы и методы, позволяющие разделять смеси, состоящие из малых количеств органических веществ. Важнейший из них – хроматография, в основе которой лежит феномен адсорбции. Разделение компонентов смеси проводят при этом либо на бумаге, либо путем адсорбции на сорбенте, которым заполняют колонки (длинные стеклянные трубки), с последующей постепенной элюцией (вымыванием) каждого из компонентов.
Разделение методом электрофореза зависит от знака и числа зарядов ионизированных молекул. Электрофорез проводят на бумаге или на каком-нибудь инертном (неактивном) носителе, таком, как крахмал, целлюлоза или каучук.
Высокочувствительный и эффективный метод разделения – газовая хроматография. Им пользуются в тех случаях, когда подлежащие разделению вещества находятся в газообразном состоянии или могут быть в него переведены.
Выделение ферментов. Последнее место в описываемом ряду – животное, орган, тканевой срез, гомогенат и фракция клеточных органелл – занимает фермент, способный катализировать определенную химическую реакцию. Выделение ферментов в очищенном виде – важный раздел в изучении метаболизма.
Сочетание перечисленных методов позволило проследить главные метаболические пути у большей части организмов (в том числе у человека), установить, где именно эти различные процессы протекают, и выяснить последовательные этапы главных метаболических путей. К настоящему времени известны тысячи отдельных биохимических реакций, изучены участвующие в них ферменты.
Клеточный метаболизм. Живая клетка – это высокоорганизованная система. В ней имеются различные структуры, а также ферменты, способные их разрушить. Содержатся в ней и крупные макромолекулы, которые могут распадаться на более мелкие компоненты в результате гидролиза (расщепления под действием воды). В клетке обычно много калия и очень мало натрия, хотя клетка существует в среде, где натрия много, а калия относительно мало, и клеточная мембрана легко проницаема для обоих ионов. Следовательно, клетка – это химическая система, весьма далекая от равновесия. Равновесие наступает только в процессе посмертного автолиза (самопереваривания под действием собственных ферментов).
Потребность в энергии. Чтобы удержать систему в состоянии, далеком от химического равновесия, требуется производить работу, а для этого необходима энергия. Получение этой энергии и выполнение этой работы – непременное условие для того, чтобы клетка оставалась в своем стационарном (нормальном) состоянии, далеком от равновесия. Одновременно в ней выполняется и иная работа, связанная со взаимодействием со средой, например: в мышечных клетках – сокращение; в нервных клетках – проведение нервного импульса; в клетках почек – образование мочи, значительно отличающейся по своему составу от плазмы крови; в специализированных клетках желудочно-кишечного тракта – синтез и выделение пищеварительных ферментов; в клетках эндокринных желез – секреция гормонов; в клетках светляков – свечение; в клетках некоторых рыб – генерирование электрических разрядов и т.д.
Источники энергии. В любом из перечисленных выше примеров непосредственным источником энергии, которую клетка использует для производства работы, служит энергия, заключенная в структуре аденозинтрифосфата (АТФ). В силу особенностей своей структуры это соединение богато энергией, и разрыв связей между его фосфатными группами может происходить таким образом, что высвобождающаяся энергия используется для производства работы. Однако энергия не может стать доступной для клетки при простом гидролитическом разрыве фосфатных связей АТФ: в этом случае она расходуется впустую, выделяясь в виде тепла. Процесс должен состоять из двух последовательных этапов, в каждом из которых участвует промежуточный продукт, обозначенный здесь X–Ф (в приведенных уравнениях X и Y означают два разных органических вещества; Ф – фосфат; АДФ – аденозиндифосфат):
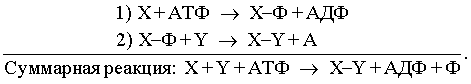
Поскольку практически для любого проявления жизнедеятельности клеток необходим АТФ, неудивительно, что метаболическая активность живых клеток направлена в первую очередь на синтез АТФ. Этой цели служат различные сложные последовательности реакций, в которых используется потенциальная химическая энергия, заключенная в молекулах углеводов и жиров (липидов).
Метаболизм углеводов и липидов
Синтез АТФ. Анаэробный (без участия кислорода). Главная роль углеводов и липидов в клеточном метаболизме состоит в том, что их расщепление на более простые соединения обеспечивает синтез АТФ. Несомненно, что те же процессы протекали и в первых, самых примитивных клетках. Однако в атмосфере, лишенной кислорода, полное окисление углеводов и жиров до CO2 было невозможно. У этих примитивных клеток имелись все же механизмы, с помощью которых перестройка структуры молекулы глюкозы обеспечивала синтез небольших количеств АТФ. Речь идет о процессах, которые у микроорганизмов называют брожением. Лучше всего изучено сбраживание глюкозы до этилового спирта и CO2 у дрожжей.
![]()
В ходе 11 последовательных реакций, необходимых для того, чтобы завершилось это превращение, образуется ряд промежуточных продуктов, представляющих собой эфиры фосфорной кислоты (фосфаты). Их фосфатная группа переносится на аденозиндифосфат (АДФ) с образованием АТФ. Чистый выход АТФ составляет 2 молекулы АТФ на каждую молекулу глюкозы, расщепленную в процессе брожения. Аналогичные процессы происходят во всех живых клетках; поскольку они поставляют необходимую для жизнедеятельности энергию, их иногда (не вполне корректно) называют анаэробным дыханием клеток.
У млекопитающих, в том числе у человека, такой процесс называется гликолизом и его конечным продуктом является молочная кислота, а не спирт и CO2. Вся последовательность реакций гликолиза, за исключением двух последних этапов, полностью идентична процессу, протекающему в дрожжевых клетках.
Аэробный (с использованием кислорода). С появлением в атмосфере кислорода, источником которого послужил, очевидно, фотосинтез растений, в ходе эволюции развился механизм, обеспечивающий полное окисление глюкозы до CO2 и воды, – аэробный процесс, в котором чистый выход АТФ составляет 38 молекул АТФ на каждую окисленную молекулу глюкозы. Этот процесс потребления клетками кислорода для образования богатых энергией соединений известен как клеточное дыхание (аэробное). В отличие от анаэробного процесса, осуществляемого ферментами цитоплазмы, окислительные процессы протекают в митохондриях. В митохондриях пировиноградная кислота – промежуточный продукт, образовавшийся в анаэробной фазе – окисляется до СО2 в шести последовательных реакциях, в каждой из которых пара электронов переносится на общий акцептор – кофермент никотинамидадениндинуклеотид (НАД). Эту последовательность реакций называют циклом трикарбоновых кислот, циклом лимонной кислоты или циклом Кребса. Из каждой молекулы глюкозы образуется 2 молекулы пировиноградной кислоты; 12 пар электронов отщепляется от молекулы глюкозы в ходе ее окисления, описываемого уравнением:
![]()
Перенос электронов. В каждой митохондрии имеется механизм, посредством которого восстановленный НАД (НАДН, где Н – водород), образовавшийся в цикле трикарбоновых кислот, передает свою пару электронов кислороду. Перенос, однако, не происходит напрямую. Электроны как бы передаются «из рук в руки» и, лишь пройдя цепь переносчиков, присоединяются к кислороду. Эта «цепь переноса электронов» состоит из следующих компонентов:
НАДНН Флавинадениндинклеотид Кофермент Q
Цитохром b Цитохром c Цитохром a O2
Все компоненты этой системы, находящиеся в митохондриях, фиксированы в пространстве и сцеплены друг с другом. Такое их состояние облегчает перенос электронов.
В состав НАД входит никотиновая кислота (витамин ниацин), а в состав флавинадениндинуклеотида – рибофлавин (витамин B2). Кофермент Q представляет собой высокомолекулярный хинон, синтезируемый в печени, а цитохромы – это три разных белка, каждый из которых, подобно гемоглобину, содержит гемогруппу.
В цепи переноса электронов на каждую пару электронов, перенесенную от НАДН на O2, синтезируется 3 молекулы АТФ. Поскольку от каждой молекулы глюкозы отщепляются и передаются молекулам НАД 12 пар электронов, в общей сложности на каждую молекулу глюкозы образуется 312 = 36 молекул АТФ. Этот процесс образования АТФ в ходе окисления называется окислительным фосфорилированием.
Липиды как источник энергии. Жирные кислоты могут использоваться в качестве источника энергии приблизительно так же, как и углеводы. Окисление жирных кислот протекает путем последовательного отщепления от молекулы жирной кислоты двууглеродного фрагмента с образованием ацетилкофермента A (ацетил-КоА) и одновременной передачей двух пар электронов в цепь переноса электронов. Образовавшийся ацетил-КоА – нормальный компонент цикла трикарбоновых кислот, и в дальнейшем его судьба не отличается от судьбы ацетил-КоА, поставляемого углеводным обменом. Таким образом, механизмы синтеза АТФ при окислении как жирных кислот, так и метаболитов глюкозы практически одинаковы.
Если организм животного получает энергию почти целиком за счет одного только окисления жирных кислот, а это бывает, например, при голодании или при сахарном диабете, то скорость образования ацетил-КоА превышает скорость его окисления в цикле трикарбоновых кислот. В этом случае лишние молекулы ацетил-КоА реагируют друг с другом, в результате чего образуются в конечном счете ацетоуксусная и -гидроксимасляная кислоты. Их накопление является причиной патологического состояния, т.н. кетоза (одного из видов ацидоза), который при тяжелом диабете может вызвать кому и смерть.
Запасание энергии. Животные питаются нерегулярно, и их организму нужно как-то запасать заключенную в пище энергию, источником которой являются поглощенные животным углеводы и жиры. Жирные кислоты могут запасаться в виде нейтральных жиров либо в печени, либо в жировой ткани. Углеводы, поступая в большом количестве, в желудочно-кишечном тракте гидролизуются до глюкозы или иных сахаров, которые затем в печени превращаются в ту же глюкозу. Здесь из глюкозы синтезируется гигантский полимер гликоген путем присоединения друг к другу остатков глюкозы с отщеплением молекул воды (число остатков глюкозы в молекулах гликогена доходит до 30 000). Когда возникает потребность в энергии, гликоген вновь распадается до глюкозы в реакции, продуктом которой является глюкозофосфат. Этот глюкозофосфат направляется на путь гликолиза – процесса, составляющего часть пути окисления глюкозы. В печени глюкозофосфат может также подвергнуться гидролизу, и образующаяся глюкоза поступает в кровоток и доставляется кровью к клеткам в разных частях тела.
Синтез липидов из углеводов. Если количество углеводов, поглощенных с пищей за один прием, больше того, какое может быть запасено в виде гликогена, то избыток углеводов превращается в жиры. Начальная последовательность реакций совпадает при этом с обычным окислительным путем, т.е. сначала из глюкозы образуется ацетил-КоА, но далее этот ацетил-КоА используется в цитоплазме клетки для синтеза длинноцепочечных жирных кислот. Процесс синтеза можно описать как обращение обычного процесса окисления жирных клеток. Затем жирные кислоты запасаются в виде нейтральных жиров (триглицеридов), отлагающихся в разных частях тела. Когда требуется энергия, нейтральные жиры подвергаются гидролизу и жирные кислоты поступают в кровь. Здесь они адсорбируются молекулами плазменных белков (альбуминов и глобулинов) и затем поглощаются клетками самых разных типов. Механизмов, способных осуществлять синтез глюкозы из жирных кислот, у животных нет, но у растений такие механизмы имеются.
Метаболизм липидов. Липиды попадают в организм главным образом в форме триглицеридов жирных кислот. В кишечнике под действием ферментов поджелудочной железы они подвергаются гидролизу, продукты которого всасываются клетками стенки кишечника. Здесь из них вновь синтезируются нейтральные жиры, которые через лимфатическую систему поступают в кровь и либо транспортируются в печень, либо отлагаются в жировой ткани. Выше уже указывалось, что жирные кислоты могут также синтезироваться заново из углеводных предшественников. Следует отметить, что, хотя в клетках млекопитающих может происходить включение одной двойной связи в молекулы длинноцепочечных жирных кислот (между С–9 и С–10), включать вторую и третью двойную связь эти клетки неспособны. Поскольку жирные кислоты с двумя и тремя двойными связями играют важную роль в метаболизме млекопитающих, они в сущности являются витаминами. Поэтому линолевую (C18:2) и линоленовую (C18:3) кислоты называют незаменимыми жирными кислотами. В то же время в клетках млекопитающих в линоленовую кислоту может включаться четвертая двойная связь и путем удлинения углеродной цепи может образоваться арахидоновая кислота (C20:4), также необходимый участник метаболических процессов.
В процессе синтеза липидов остатки жирных кислот, связанные с коферментом А (ацил-КоА), переносятся на глицерофосфат – эфир фосфорной кислоты и глицерина. В результате образуется фосфатидная кислота – соединение, в котором одна гидроксильная группа глицерина этерифицирована фосфорной кислотой, а две группы – жирными кислотами. При образовании нейтральных жиров фосфорная кислота удаляется путем гидролиза, и ее место занимает третья жирная кислота в результате реакции с ацил-КоА. Кофермент А образуется из пантотеновой кислоты (одного из витаминов). В его молекуле имеется сульфгидрильная (– SH) группа, способная реагировать с кислотами с образованием тиоэфиров. При образовании фосфолипидов фосфатидная кислота реагирует непосредственно с активированным производным одного из азотистых оснований, таких, как холин, этаноламин или серин.
За исключением витамина D, все встречающиеся в организме животных стероиды (производные сложных спиртов) легко синтезируются самим организмом. Сюда относятся
8-09-2015, 19:25