Слуховая система: кодирование частоты звука
Слуховая система: кодирование частоты звука
Соматосенсорная система анализирует стимулы на основе их расположения на карте тела. Благодаря аналогичному процессу, зрительные стимулы являются ретинотопически картированными и анализируются на предмет их местоположения в визуальном (пространственном) мире. Однако слуховая система использует совершенно иной метод анализа. Слуховая периферия, или улитка, картирует звуковые стимулы скорее по их частотному содержанию, чем по пространственному расположению, и последующие стадии слуховой системы используют этот тонотопический анализ, чтобы определить смысл речи и значение других сложных звуков. Как частота звука кодируется волосковыми клетками улитки, и как эти сигналы обрабатываются на более высоких уровнях слуховой системы? Как определяется пространственное расположение звука?
У наземных позвоночных звуковые волны в воздухе входят во внешнее ухо, колеблют барабанную перепонку и, благодаря серии механических сопряжений в среднем ухе, конвертируются в колебания жидкости улитки. Колебания жидкости, в свою очередь, вызывают вибрацию базилярной мембраны, на которой находятся сенсорные волосковые клетки в составе органа Корти. Этот процесс описан поэтически Олдосом Хаксли:
Трубный звук Понгилеони и скрежет безымянных скрипок потрясли воздух в большой зале, вызвали дрожь в оконных стеклах, и это, в свою очередь, сотрясло воздух в апартаментах лорда Эдварда на дальней стороне. Сотрясающийся воздух задребезжал в барабанных перепонках лорда Эдварда; соединенные между собой косточки молоточка (ушной косточки), наковальни внутреннего уха и стремени (слуховой кости среднего уха) были приведены в движение, так что возбудили мембрану окна (oval window) и подняли бесконечно малую бурю в жидкости лабиринта. Волосковые окончания слухового нерва дрожали, как водоросли в бушующем море; огромное количество непонятных чудес произошло в мозгу, и лорд Эдвард в экстазе прошептал: «Бах!»
Улитка
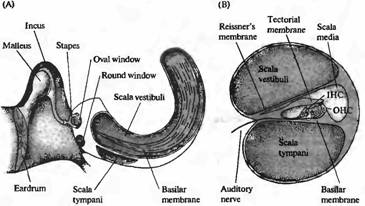
Канал улитки разделен на три отдела. Средняя лестница (scala media) содержит раствор с высоким содержанием калия, эндолимфу. Он отделен от покрывающей его вестибулярной лестницы (scala vestibuli) Рейсснеровой мембраной, а от наполненного жидкостью пространства барабанной лестницы (scala timpani) — тесными межклеточными соединениями между апикальными окончаниями волосковых клеток и окружающими вспомогательными клетками (рис. 1В). Барабанная и вестибулярная лестницы содержат перилимфу, сходную по составу с цереброспинальной жидкостью; средняя лестница содержит эндолимфу, ионный состав которой похож на состав цитоплазмы: высокое содержание калия, низкое — натрия и с концентрацией кальция, поддерживаемой на микромолярном уровне. Необычный состав внеклеточной жидкости устанавливается благодаря работе ионных насосов в клетках, расположенных в сосудистом слое (stria vascularis), т. е. в секреторном эпителии, выстилающем боковую стенку средней лестницы.
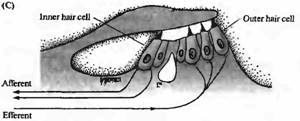
Существуют две различные группы волосковых клеток в улитке млекопитающих: внутренние волосковые клетки и внешние волосковые клетки. Они различаются по своему расположению (внутренние волосковые клетки ближе к центральной оси спирали улитки) и по паттерну иннервации. Внутренние волосковые клетки получают более 90% афферентных контактов улитки; внешние волосковые клетки являются постсинаптическими мишенями для эфферентной иннервации (рис. 1С). Эти различия в паттернах иннервации ставят интересные вопросы, касающиеся функциональной роли этих двух типов клеток, которые будут обсуждаться далее в этой главе. Покровная мембрана (tectorial membrane) налегает на волосковые пучки как внутренних, так и внешних волосковых клеток, и различие в движении покровной и базилярной мембран приводит к боковому сдвигу, открывающему каналы механотрансдукции.
Частотная избирательность: механическая настройка
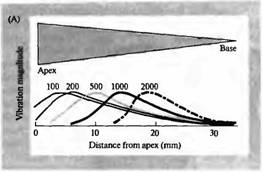
Звуковое восприятие — это не просто вопрос выявления звукового стимула. Информационное содержание звука является функцией его частотного состава. Таким образом, слуховая система зависит от способности ее механосенсорных волосковых клеток быть частотно избирательными. В улитке млекопитающих настройка по частоте зависит от механических свойств специализированной структуры, в которой находятся сенсорные клетки, аналогично механической адаптации, осуществляемой многослойной капсулой в тельце Пачини. Ширина и толщина базилярной мембраны закономерно меняется по длине канала улитки (рис. 1 А). У основания улитки (ближе к овальному окну) базилярная мембрана узкая и жесткая; у противоположного конца (вершина улитки) она широкая и гибкая. По причине этой изменяющейся жесткости входящие звуковые волны вызывают движение жидкости и вибрацию базилярной мембраны в разных участках в зависимости от частоты звука.
Это свойство функции улитки было описано фон Бекеси, который использовал стробоскопическое освещение отражающих частиц, рассеянных в улитке, чтобы визуализировать паттерн вибрации. Он обнаружил, что звуки высокой частоты вызывают максимальные колебания на более толстой, жест кой нижней части мембраны, а звуки низкой частоты вызывают максимальную вибрацию на более гибкой верхней части мембраны улитки. Фон Бекеси назвал этот паттерн колебаний «бегущей волной».
Как следствие механической настройки базилярной мембраны,
|
Рис. 1. Структура улитки. (А) Схема среднего уха и улитки, показывающая систему косточек, с помощью которых барабанная перепонка соединена с мембраной овального окна улитки. Улитка дана рассеченной и не скрученной, чтобы показать ее основные внутренние полости (вестибулярную и барабанную лестницы) и форму базилярной мембраны, расширяющейся от основания улитки к ее вершине. (В) Поперечный разрез улитки, показывающий структурные взаимоотношения между базилярной мембраной, внутренними (IHC) и наружными (ОНС) волосковыми клетками и покровной мембраной. (С) Иннервация волосковых клеток улитки. Около 95 % афферентных волокон являются постсинаптическими мишенями для IHC. До 20 афферентных волокон контактирует с одиночной IHC. ОНС имеют мало афферентных связей, зато являются постсинаптическими мишенями для холинергических эфферентных нейронов, которые проецируются из оливарного комплекса, расположенного в стволе мозга. |
волосковые клетки и их постсинаптические афферентные волокна у основания улитки преимущественно стимулируются высокочастотным звуком, тогда как те, что находятся у вершины улитки, лучше всего реагируют на низкочастотный звук (рис. 2). Частотная избирательность афферентов улитки может быть измерена при помощи отведения потенциалов действия, возникающих в ответ на чистые тона звука,
Рис. 2. Настройка улитки. (А) Локализация участков улитки, в которых звуковые волны вызывают максимальное смещение базилярной мембраны, зависит от частоты. Кривые представляют относительное смещение на указанных частотах (100-2000 Гц). (В) Типичные кривые настройки четырех индивидуальных волокон слухового нерва, иннервирующих различные участки улитки. Показана зависимость между частотой звукового стимула и интенсивностью звука в децибелах (dB) необходимой для возникновения разряда в волокне. Пики соответствуют оптимальным звуковым частотам (т. е. частотам, при которых интенсивность стимула для каждого волокна минимальна). against frequency of the auditory stimulus. |
|
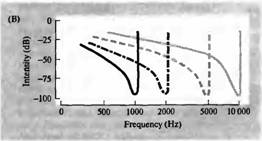
частота и интенсивность которых систематически варьирует. Результирующая кривая настройки V-образна, с оптимальной, или характеристической, частотой, определяемой как чистый тон, к которому волокно наиболее чувствительно. Характеристическая частота каждого волокна определяется по тому, где именно в канале улитки оно контактирует с волосковой клеткой.
Звуковая частота картируется на протяжении эпителиального массива рецепторов, производящих кохлеотопическую, или тонотопическую карту частоты. Поэтому, слуховая система может использовать принципы нейронной обработки — такие, как латеральное торможение — основанные на тонотопии, чтобы истолковывать и обрабатывать информацию, закодированную в звуковой частоте таким же образом, как соматотопические карты лежат в основе обработки соматосенсорных сигналов.
Эфферентное торможение улитки
Как и мышечные веретена, слуховые рецепторы подвергаются эфферентной регуляции. Нейроны комплекса верхних олив (olivary complex) в стволе мозга млекопитающих проецируются на ипсилатералъную и контралатеральную улитку. Активация этого пути вызывает высвобождение ацетилхолина в эфферентных синапсах на волосковых клетках и подавляет ответы на звук в афферентных волокнах улитки. Эфферентная обратная связь снижает чувствительность улитки при наличии фонового шума и снижает риск перегрузки. Это — аналог переустановки чувствительности мышечного веретена гамма-эфферентами, поддерживающими поток информации во время движения конечности в широком диапазоне положений. Эфферентные волокна активируются звуком и иннервируют ограниченные области улитки; в результате подавление шума носит частотно-специфический характер. В дополнение к восстановлению динамического диапазона торможение расширяет кривую настройки афферентных волокон. Наконец, эфферентная обратная связь может защитить улитку от повреждения громким звуком. Фактически, сила эфферентной обратной связи находится в обратном соотношении со степенью акустического повреждения, вызванного громким звуком.
Активация эфферентного пути к уху черепахи вызывает большие гиперполяризирующие тормозные постсинаптические потенциалы (IPSP) в волосковых клетках. Влияние торможения на ответ волосковой клетки на звук показан на рис. 3А. Клетка была стимулирована чистыми тонами на трех частотах, одна из которых является оптимальной для данной болосковой клетки, вторая — более высокой частотой, и третья — более низкой частотой. Интенсивности тонов были отрегулированы так, что все они вызывали осциллирующий рецепторный потенциал одной и той же амплитуды. Короткая серия стимулов, поданная на эфферентное волокно, гиперполяризировала клетку и сильно ослабила ее ответ на тон при 220 Гц (характеристическая частота).
Рис. 3. Влияние эфферентной стимуляции волосковых клеток улитки на звуковые стимулы. (А) Средняя запись: осцилляторный ответ волосковой клетки на звуковой стимул частотой 220 Гц (резонансная частота) тормозится короткой пачкой эфферентных стимулов (обозначено полоской), и клетка гиперполяризуется. Верхняя запись: ответ на звуковой стимул при 70 Гц. Интенсивность стимула подобрана таким образом, что осцилляторный ответ был близок по амплитуде ответу на звуковой стимул частотой 220 Гц. Эфферентная стимуляция также вызывает гиперполяризацию, однако не ослабляет, а, наоборот, усиливает осцилляторный ответ. Нижняя запись: то же при частоте 857 Гц, однако этот ответ не изменяется при эфферентной стимуляции. (В) Чувствительность другой волосковой клетки (в мВ на единицу звукового давления) показана как функция частоты при эфферентной стимуляции (серым цветом) и в ее отсутствии (черным). Эфферентное торможение снижает ответ при резонансной частоте и повышает чувствительность на более низких частотах, в результате чего настройка нарушается. |
|
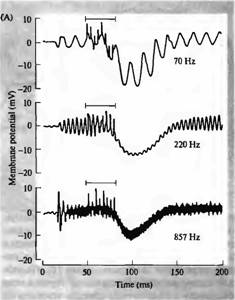
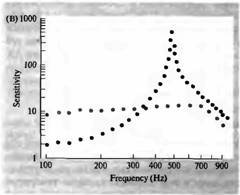
На более низкой и высокой частотах акустической стимуляции активация эфферентов по-прежнему приводила к гиперполяризации, но низкочастотная осцилляция даже увеличивалась по амплитуде, тогда как на высоких частотах амплитуда была неизменна. Этот дифференциальный эффект торможения приводит к расширению частотного диапазона ответов волосковых клеток (рис. 3В).
Механизм холинергического торможения был исследован в волосковых клетках цыпленка. Ацетилхолиновые (АХ) рецепторы
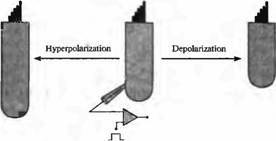 |
Рис. 4. Электрическая подвижность наружных вол сковых клеток. Для фиксации мембранного потенциала наружных волосковых клеток, изолированных из улитки позвоночных, были использованы пэтч пипетки Деполяризация приводила к укорочению клеток; гилерполяризация делала их длиннее. Такие изменения длины могли быть достаточно веяики, достигая 30 нм/мВ. |
являются лиганд-регулируемыми катионными каналами, через которые натрий и кальций входят в клетку, приводя к активации кальцийзависимых калиевых каналов и к гиперполяризации мембраны. Подобный холинергический механизм торможения обнаружен в волосковых клетках млекопитающих. Ответы волосковых клеток на АХ блокируются бунгаротоксином, и есть надежное доказательство того, что необычный член семейства никотиновых рецепторов является лиганд-связующей субъединицей АХ рецептора волосковой клетки.
Электрическая подвижность волосковых клеток улитки млекопитающих
Эфферентные нейроны, которые тормозят ответы улитки, образуют синапсы на внешних волосковых клетках, но не на внутренних волосковых клетках, которые генерируют нейрональный выход улитки млекопитающих. Как этот тормозящий эффект передается от внешней к внутренней волосковой клетке? Важное свойство внешней волосковой клетки в улитке млекопитающих — электрическая подвижность. Внешние волосковые клетки, изолированные из улитки млекопитающих, укорачиваются, реагируя на деполяризацию, и удлиняются во время гиперполяризации их мембран (рис. 4). Эти движения не вызываются актин-миозином, но возникают в результате прямых эффектов электрического потенциала на заряженные «моторные» протеины в базолатеральной мембране. Каков бы ни был механизм электрической подвижности внешних волосковых клеток, она также способствует вибрации структуры улитки во время звуковой стимуляции, и, таким образом, увеличивает отклонение стереоцилиев внутренних волосковых клеток.
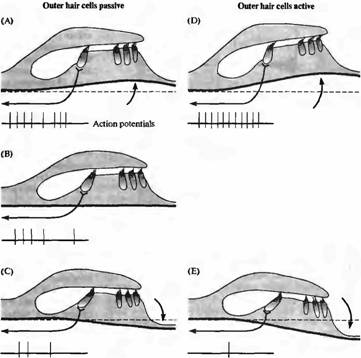
Внешние волосковые клетки улучшают настройку улитки, сообщая дополнительную механическую энергию движению базилярной мембраны (рис. 5). Это объясняет, как эфферентные синапсы, образованные на внешних волосковых клетках, подавляют звуковой ответ внутренних волосковых клеток. Поскольку подвижность внешних волосковых клеток вызывается изменениями мембранного потенциала, синаптическое торможение действует так, что подавляет электрическую подвижность аналогично тормозным эффектам в других областях нервной системы. Торможение уменьшает активный механический вклад внешних клеток в колебания структур улитки и, таким образом, опосредованно снижает возбуждение внутренних волосковых клеток.
Электрическая настройка волосковых клеток
Уши немлекопитающих позвоночных также частотно избирательны, несмотря на то, что у них базилярные мембраны не расширяются и не поддерживают механически настроенные бегущие волны. Кроуфорд и Феттиплейс предприняли исследования на черепахе, чтобы показать, что механосенсорные волосковые клетки обладают встроенной частотной избирательностью благодаря механизму электрической настройки. Внутриклеточные отведения от волосковой клетки базилярного сосочка черепахи (слуховой сенсорный эпителий) показаны на рис. 6. Когда подавался короткий звуковой стимул (щелчок), возникали затухающие колебания мембранного потенциала волосковой клетки или осциллирующий
| Рис. 5. Движение структур улитки. Во время волновых колебаний жидкости, вызванных звуком, подвижные структуры улитки (базилярная мембрана и орган Корти) перемещаются вверх и вниз. (А) Во время фазы разрежения звуковой волны базилярная и покровная мембраны смещаются вверх, приводя к отклонению волосковых пучков в направлении наиболее длинного стереоцилия — деполяризация волосковых клеток по сравнению с состоянием покоя (В). (С) Перемещение вниз во время фазы сжатия звуковой волны вызывает противоположное отклонение волосковых пучков и волосковые клетки гиперполяриэуются относительно спокойного состояния. (D) Активное сокращение наружных волосковых клеток во время деполяризации усиливает направленное вверх смещение базилярной мембраны. (Е) Гиперполяризация наружных волосковых клеток приводит к их удлинению. Результирующий эффект состоит в увеличении смещения базилярной мембраны за счет «мышечного» действия наружных |
волосковых клеток, что отражается в паттерне активности волокон, контактирующих с наружными волосковыми клетками. |
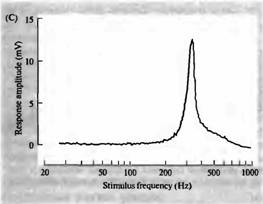
ответ с частотой около 350 Гц. Это та же частота, что и у чистых тонов, к которым волосковые клетки наиболее чувствительны, как показано на рис. 6С. Здесь тон постоянной интенсивности подавался на внешнее ухо, и его частота постепенно менялась от 20 до 1 000 Гц. Электрический ответ волосковой клетки достигал пикового значения при 350 Гц. Осцилляция потенциала, вызванная коротким звуковым сигналом, и кривая настройки, полученная путем подбора частоты, являются эквивалентными средствами для измерения частотной избирательности волосковой клетки.
Рис. 6В показывает, что когда микроэлектрод использовали для пропускания в клетку прямоугольного толчка тока, возникала осцилляция потенциала, частота и скорость затухания которой были идентичны той, что была вызвана звуковым входом. Вывод из этих экспериментов состоит в том, что частотная избирательность волосковых клеток зависит от электрических свойств мембраны. Электрически и акустически настраиваемые частоты волосковых клеток эквивалентны и закономерно изменяются по длине базилярного сосочка черепахи, создавая тонотопический массив настроенных детекторов.
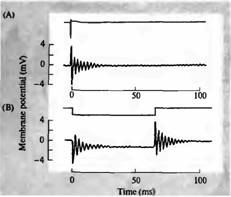 |
Рис. 6. Настройка волосковых клеток в улитке черепахи. (А) Влияние звукового щелчка (верхняя линия) на мембранный потенциал волосковой клетки (нижняя линия), зарегистрированный внутриклеточно. Щелчок вызывает затухающее колебание мембранного потенциала с частотой около 350 Гц, начальной пиковой амплитудой около 8 мВ (В) Гиперполяризующий толчок тока (верхняя линия), приложенный к той же клетке, вызывает сходные осцилляции как при включении, так и при выключении толчка, показывая, что частота колебаний является внутренним свойством волосковой клетки. (С) Если такую волосковую клетку стимулировать чистыми тонами, плавно меняющимися по частоте от 25 до 1000 Гц, пиковая амплитуда рецепторного потенциала имеет острый максимум при 350 Гц. |
 |
Калиевые каналы волосковых клеток и их настройка
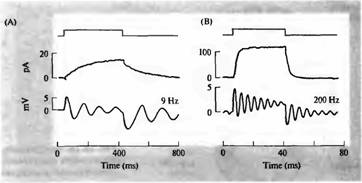
Какие свойства клеточной мембраны обеспечивают эту электрическую настройку и как эти свойства варьируют, определяя настройку на определенную частоту? Отведение от волосковых клеток мешочка лягушки (вестибулярный сенсорный орган) показало, что взаимодействия между потенциалзависимыми кальциевыми и кальций-активируемыми калиевыми каналами может вызывать осцилляцию потенциала. Исследования волосковых клеток, изолированных из уха черепахи, показали, что характеристическая частота каждой клетки определена чрезвычайно элегантным и простым путем — а именно плотностью и кинетическими свойствами кальций-активируемых калиевых каналов в каждой клетке (рис. 7). Их называют ВК-каналами (от "big K+ ") из-за большой проводимости одиночного канала.
В клетках, настроенных на более низкие частоты, суммарная калиевая проводимость мала и медленнее активируется, и таким образом способствует возникновению относительно медленных электрических колебаний. В клетках с более высокой частотой калиевая проводимость больше и активация быстрее.
В самых низкочастотных волосковых клетках еще более медленные, исключительно потенциалзависимые калиевые каналы поддерживают колебания.
ВК-каналы в волосковых клетках цыплят и черепах кодируются геном, тРНК которого обладает альтернативным сплайсингом, и некоторые дифференциально расщепляемые изоформы канала кинетически различны. Дополнительная вариабельность обеспечивается вспомогательной субъединицей, которая встраивается в канал и замедляет кинетику его открывания) .
| Рис. 7. Частота настройки и калиевая проводимость в изолированной волосковой клетке черепахи, измеренные с помощью пзтч-кламп отведения от целой клетки. (А) Средняя запись показывает выходящий ток, обусловленный, в основном, калием, возникающий в ответ на деполяризующее смещение потенциала (длительность которого показана на верхней записи). Ток медленно нарастает до максимума в 15 пА с постоянной времени около 200 мс. Небольшой сдвиг тока такой же длительности вызывает осцилляторное изменение напряжения в начале и в конце толчка (нижняя запись) с резонансной частотой 9 Гц. (В) В другой клетке сходный деполяризующий толчок вызывает значительно |
больший по величине, быстро нарастающий выходящий ток, свидетельствующий о более высокой плотности калиевых каналов с более быстрой кинетикой. Частота настройки осцилляторного ответа также пропорционально возрастает до 200 Гц. |
Эта -субъединица экспрессируется в тонотопическом паттерне, появляясь в больших количествах в клетках вершины улитки, где обнаруживается
8-09-2015, 19:43