Слуховые проводящие пути
Основные слуховые пути схематично представлены на рис. 8. Слуховые волокна восьмой пары черепномозговых нервов направляются центрально и посылают ветви как в дорзальные, так и в вентральные кохлеарные ядра. Аксоны второго порядка восходят в составе контралатерального бокового леминиска, иннервируя клетки в нижних бугорках четверохолмия (ядро бокового лемниска является станцией синаптического переключения для некоторых из этих волокон). Нейроны в вентральном кохлеарном ядре также дают коллатеральные ветви как к ипсилатеральным, так и к контралатеральным ядрам верхних олив. Клетки третьего порядка, расположенные в ядрах верхних олив, в свою очередь, посылают восходящие волокна в нижние бугры четверохолмия. Путь продолжается через ядра медиального коленчатого тела таламуса к слуховой области поперечной височной извилины верхней части височных долей коры головного мозга. Каждый уровень в слуховом пути тонотопически картируется.
По мере продвижения к более высоким отделам слуховой системы чистые тона становятся все менее важными в качестве стимулов для отдельных клеток. Например, клетки в дорзальном кохлеарном ядре и нижних буграх четверохолмия тормозятся тонами, которые не совпадают с их характеристической частотой. Клетки в слуховой коре реагируют на комбинации тонов, на бинауральные, но не мензуральные звуки, а также на другие сложные слуховые стимулы. Когда обнаруживаются клетки, которые действительно реагируют на чистые тона, их частотная избирательность зачастую выше, чем у первичных афферентных волокон. Таким образом, кривая настройки (отношение порога к частоте) корковых нейронов острее, чем кривая первичного афферента (рис.9А). Это обострение частотной избирательности ответа соответствует обострению пространственной локализации колебаний на базилярной мембране и обеспечивается за счет
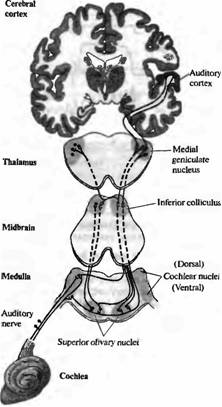 |
Рис. 8. Центральные слуховые пути. Волокна слухового нерва оканчиваются билатерально в дорзальных и вентральных кохлеарных ядрах. Волокна второго порядка восходят контралатерально к нижним буграм четверохолмия; волокна от вентральных кохлеарных ядер также дают коллатерали билатерально в ядра верхних олив. Следующее билатеральное взаимодействие обеспечивается на уровне нижних бугров четверохолмия. Затем волокна восходят к ядрам медиального коленчатого тела таламуса, а оттуда направляются в слуховую кору. |
латерального торможения, точно так же как и в соматосенсорной системе. Таким образом, рецептивным полем этого коркового нейрона является возбужденная полоса базилярной мембраны, окруженная с обеих сторон тормозным окаймлением. Это приводит к сужению диапазона частот, которые возбуждают клетку, за счет сжатия его с обеих сторон более высокими и более низкими частотами, которые вызывают торможение (рис. 9В). Тормозные боковые полосы частот обусловлены частично нисходящей обратной связью из самой коры, которая формирует свойства ответа клеток, расположенных на более низких уровнях.
Литература
1.Brain Res. Rev. 26: 230-235.
2. Nathans, J.. The genes for color vision. Sci. Am. 260(2): 42-49.
3. Sterling, P. Retina. In GShepherd (éd.), Synoptic Organization of the Brain. Oxford University Press, New York, Chapter 6.
4. Stryer, L. 1991-1992. Molecular mechanism of visual excitation. Harvey Lect. 87: 129-143.
8-09-2015, 19:43