Санкт-Петербургская Государственная Педиатрическая Медицинская Академия
Реферат
по патологической физиологии
тема:
«Хроническая боль как типовой патологический процесс»
выполнил: студент 437 гр.
Кашин А.С.
Москва
2009
Введение
Несмотря на постоянный рост числа анальгезирующих фармацевтических средств, несмотря на увеличение физиотерапевтических, нейрохирургических, психотерапевтических методов лечения, несмотря на широкое распространение нейровизуальных и нейрофизиологических способов выявления скрытых и латентно протекающих органических поражений, эффективность лечения пациентов, страдающим различными видами болевых синдромов остаётся очень низкой. Это приводит к частому и неоправданному использованию сильнодействующих и наркотических препаратов, к возникновению опасных лекарственных осложнений и аддикций, ставит в трудное положение медиков, как перед своими пациентами, так и перед правоохранительными органами, контролирующими борьбу с наркоманиями.
Основные направления решения этой проблемы состоят, во-первых, в проведении чёткой границы между острой, физиологической, и хронической, патологической, болью, во-вторых, в необходимости изменения концептуального подхода к понятию патологической (хронической) боли, не как симптому болезни, а как самостоятельному патологическому процессу, либо заболеванию.
Физиологическая боль
В историческом развитии теорий болевой перцепции можно выделить 3 основных этапа (R. Melzack, 1993).
Первый из них был связан с именем французского философа и физиолога Р. Декарта. Его учение о рефлексе послужило основой для создания в последующем "теории специфичности", согласно которой боль представляет собой отдельную сенсорную систему, в которой любой повреждающий стимул активирует специальные болевые рецепторы (ноцицепторы), передающие болевой импульс по специальным нервным путям в спинной мозг и в болевые центры головного мозга, вызывая ответную защитную реакцию, направленную на удаление от раздражителя.
К середине 20-го столетия правомерность концепции боли, как специфической проекционной сенсорной системы, была подтверждена многочисленными исследованиями и открытиями в анатомии и экспериментальной физиологии. Были обнаружены болепроводящие нервные волокна и болепроводящие пути в спинном мозге, болевые центры в различных отделах головного мозга.
Согласно теории специфичности, психологическое ощущение боли, её восприятие и переживание признаются адекватными и пропорциональными физической травме и периферическому повреждению.
В практической медицинской деятельности это положение привело к тому, что пациенты, страдающие болью и не имеющие очевидных признаков органической патологии стали считаться "ипохондриками", "невротиками" и, в лучшем случае, направлялись на лечение к психиатру или психотерапевту.
Клиническим применением теории специфичности стало широкое распространение в 50х годах 20-го века различных нейрохирургических методов, разрушающих болепроводящие пути или болевоспринимающие центры (невротомия, ризотомия, хордотомия и др.). Однако изучение отдалённых результатов этих операций показало, что применение этих методов к больным, страдающим хронической и патологической болью, приносит лишь кратковременный эффект. Примером может служить гидротермическая деструкция ветвей тройничного нерва, неизбежно приводящая к рецидиву болей в ещё более жестоком варианте.
Поэтому многие специалисты прекрасно понимали несовершенство теории специфичности, отводившей центральной нервной системе роль пассивного приёмника болевых импульсов. С 19-го века предпринимались многочисленные попытки создать новую теорию боли, основанную на суммации нервных раздражений до определённого порога, за которым возникает болевое ощущение. "Теория паттернов" имела несколько вариантов, эволюционировавших в 60-е годы в теорию "воротного контроля" (R. Melzack, P. Wall, 1965).
Основные положения теории "воротного контроля" заключались в следующем (рис. 1.):
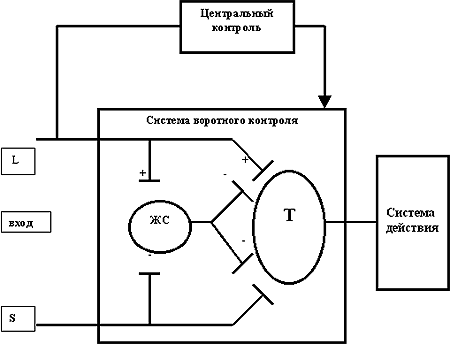
ЖС - желатинозная субстанция задних рогов спинного мозгаТ - трансмиссивные нейроныL - волокна большого диаметраS - волокна малого диаметра
Рис. 1. Схема воротного механизма контроля боли (R.Melzack, 1999)
1. Передача нервных импульсов в центральную нервную систему модулируется специальными "воротными" механизмами, расположенными в задних рогах спинного мозга.
2. Спинальные воротные механизмы представляют собой взаимосвязь активности афферентных волокон большого диаметра (L) и волокон малого диаметра (S): активность L-волокон тормозит передачу импульсов ("закрывает ворота"), в то время как активность S-волокон облегчает их передачу ("открывает ворота").
3. Спинальные "воротные" механизмы, в свою очередь, также регулируются нисходящими импульсами от головного мозга, активируемыми системой быстропроводящих волокон большого диаметра (L).
4. При достижении критического уровня поток импульсов от нейронов спинного мозга (релейные, трансмиссивные нейроны, передаточные Т-клетки) активирует Систему действия, т.е. те нейрональные зоны центральной нервной системы, которые формируют сложные поведенческие реакции на боль.
Следует заметить, что только в середине 70-х годов эта теория получила широкое признание и была опубликована во всех ведущих руководствах по медицине.
Теория "воротного контроля" имела огромное клиническое значение. Методы перерезки нервов и болепроводящих путей стали постепенно замещаться методами воздействия на входящую в спинной мозг информацию. Физиотерапевты, рефлексотерапевты и специалисты в области лечебной гимнастики, использующие множество модулирующих методик, включая акупунктуру и чрезкожную электронейростимуляцию (ЧЭНС), заняли важное место в терапии острой и хронической боли.
Основное научно-медицинское значение теории "входных ворот" заключалось в признании спинного и головного мозга активной системой, фильтрующей, отбирающей и воздействующей на входные сенсорные сигналы. Теория утвердила центральную нервную систему ведущим звеном в болевых процессах.
Однако с течением времени появились новые факты, которые невозможно было объяснить даже с позиции этой теории. И, прежде всего, они касались наблюдений за больными с параплегией, т.е. с перерывом спинного мозга, и пациентами, страдающими фантомными болями, сохраняющих переживание и ощущение уже отсутствующих частей тела (R.Melzack, 1989).
Анализ феномена фантомных болей привёл к следующим заключениям:
во-первых, поскольку фантомная конечность ощущается настолько реальной, то следует, что её нормальное ощущение обусловлено процессами в самом головном мозге, а, значит, может возникать и в отсутствие входных проприоцептивных сигналов;
во-вторых, поскольку все сенсорные ощущения, включая боль, также могут возникать в отсутствие раздражителей, можно считать, что источники возникновения нервных паттернов, формирующих качество переживания, находятся не в периферической нервной системе, а в нейрональных сетях головного мозга.
Следовательно, восприятие собственного тела и его многообразных ощущений обусловлено центральными процессами в головном мозге, генетически детерминировано и может лишь модифицироваться под воздействием периферических сигналов и прошлого опыта.
Этот вывод стал основой теории, утверждавшей новую концептуальную модель нервной системы, теории нейроматрикса (R. Melzack, 1999).
Нейроматрикс представляет собой обширную сеть нейронов, образующих функциональные петли между таламусом и корой, корой и лимбической системой. Синаптические связи в этой нейронной сети генетически детерминированы и, в некотором смысле, составляют материнскую "матрицу", генерирующую, воспроизводящую и модулирующую сенсорную информацию.
Нейронные петли дивергируют, т.е. расходятся в пространстве и формируют три основных компонента нейроматрикса - "нейромодули", в которых ведётся одновременная параллельная обработка данных. Нейромодули соответствуют трём главным психологическим компонентам болевого ощущения: сенсорно-дискриминативному, аффективно-мотивационному и оценочно-познавательному.
Сенсорно-дискриминативные процессы позволяют оценить качество, силу, длительность болевого воздействия. Они зависят и определяются потоком импульсов, возникающих вследствие раздражения рецепторов кожи, мышц, висцеральных органов и других соматических тканей.
Аффективно-мотивационный компонент формирует потребность в действиях, направленных на то, чтобы избежать боли. Он зависит от фоновых, тонических свойств нервной системы, определяющихся культурно-этическими традициями, уровнем образования, личностными характеристиками и др.
Оценочно-познавательный процесс восприятия боли обеспечивается фазическими, временными параметрами нервной деятельности, - уровнем внимания, тревожности, слуховым и зрительным участием, памятью и опытом.
Обработка данных в нейромодулях завершается конвергенцией (схождением) информации по нейронным петлям, взаимодействием между ними и формированием конечного результата - созданием характерного паттерна сигналов - "нейросигнатуры".
Нейросигнатура представляет собой непрерывный, исходящий из нейроматрикса поток (паттерн) генетически детерминированных и ситуационно изменённых импульсов, направляющихся в следующий компонент системы - "сенсорный невральный центр", в котором происходит оценка этого потока сигналов, его осознание, осмысление и накопление опыта.
Одновременно такой же паттерн сигналов (нейросигнатур) направляется в нейроматрикс действия и далее в спинной мозг, вызывая мышечные реакции для выполнения простых и быстрых ответных движений и действий.
В нейроматриксе действия и сенсорном невральном центре происходит одновременная и параллельная циклическая обработка нескольких возможных вариантов реагирования и поведения, последовательное устранение ненужных и выбор наиболее соответствующего конкретным специфическим обстоятельствам варианта.
Таким образом, хотя нейроматрикс и предопределён генетическими факторами, его синаптическая индивидуальная архитектура формируется и определяется поступающими в него в течение жизнедеятельности человека сенсорными сигналами и воздействиями. Нейроматрикс представляет собой неразделимое единство наследственности, опыта и обучения.
Теория нейроматрикса утверждает, что все качественные характеристики болевого ощущения генетически детерминированы и генерируются в головном мозге, а периферические стимулы представляют лишь их неспецифические "триггеры".
Согласно новой концепции головной мозг не только воспринимает, анализирует и модулирует входные сенсорные сигналы. Он обладает свойством генерировать болевую перцепцию даже в случаях, когда никакие внешние импульсы и раздражения с периферии не поступают.
Теория нейроматрикса вероятно будет иметь значительную клиническую ценность в лечении упорных, в частности, фантомных болей. Так, например, введение местного анестетика (лидокаина) в определённые зоны головного мозга (боковой гипоталамус, зубчатое ядро и др.), что делается достаточно легко и безопасно, может блокировать процесс формирования болевых нейросигнатур на период, значительно более длительный, чем продолжительность фармакологического действия препарата (R.Melzack, 1993).
Антиноцицептивная система
Комплекс ноцицептивной системы в равной степени сбалансирован в организме комплексом антиноцицептивной системы, обеспечивающей контроль за активностью структур, участвующих в восприятии, проведении и анализе болевых сигналов.
В настоящее время установлено, что болевые сигналы, поступающие с периферии, стимулируют активность различных отделов центральной нервной системы (околопроводное серое вещество, ядра шва ствола мозга, ядра ретикулярной формации, ядра таламуса, внутренней капсулы, мозжечка, интернейроны задних рогов спинного мозга и др.) оказывающих нисходящее тормозное действие на передачу ноцицептивной афферентации в дорзальных рогах спинного мозга.
В механизмах развития анальгезии наибольшее значение придаётся серотонинергической, норадренергической, ГАМКергической и опиоидергической системам мозга (М.Л. Кукушкин, В.К. Решетняк, 1997).
Основная из них, опиоидергическая система, образована нейронами, тело и отростки которых содержат опиоидные пептиды (бета-эндорфин, мет-энкефалин, лей-энкефалин, динорфин).
Связываясь с определёнными группами специфических опиоидных рецепторов (мю-, дельта- и каппа-опиоидные рецепторы), 90% которых расположено в дорзальных рогах спинного мозга, они способствуют высвобождению различных химических веществ (гамма-аминомасляная кислота), тормозящих передачу болевых импульсов.
Эта природная, естественная болеутоляющая система так же важна для нормальной жизнедеятельности, как и болесигнализирующая система. Благодаря ей, незначительные повреждения типа ушиба пальца или растяжения связок вызывают сильные болевые ощущения только на короткое время - от несколько минут до нескольких часов, не заставляя нас страдать в течение дней и недель, что случилось бы в условиях сохранения боли до полного заживления (R. Melzack, 1999).
Таким образом, физиологическая ноцицепция включает четыре основных процесса (Н. Катц, Ф.М. Ферранте, 1998):
1. Трансдукцию - процесс, при котором повреждающее воздействие трансформируется в виде электрической активности в свободных неинкапсулированных нервных окончаниях (ноцицепторах). Их активация происходит либо вследствие прямых механических или термических стимулов, либо под воздействием эндогенных тканевых и плазменных алгогенов, образующихся при травме или воспалении (гистамин, серотонин, простагландины, простациклины, цитокины, ионы К+ и Н+, брадикинин).
2. Трансмиссию - проведение возникших импульсов по системе чувствительных нервных волокон и путей в центральную нервную систему (тонкие миелиновые А-дельта и тонкие безмиелиновые С-афференты в составе аксонов спинномозговых ганглиев и задних спинномозговых корешков, спиноталамические, спиномезенцефалические и спиноретикулярные пути, идущие от нейронов задних рогов спинного мозга к образованиям таламуса и лимбико-ретикулярного комплекса, таламокортикальные пути к соматосенсорным и фронтальной зонам коры головного мозга).
3. Модуляцию - процесс изменения ноцицептивной информации нисходящими, антиноцицептивными влияниями центральной нервной системы, мишенью которых являются преимущественно нейроны задних рогов спинного мозга (опиоидергические и моноаминовые нейрохимические антиноцицептивные системы и система воротного контроля).
4. Перцепцию - субъективное эмоциональное ощущение, воспринимаемое как боль и формирующееся под воздействием фоновых генетически детерминированных свойств центральной нервной системы и ситуационно меняющихся раздражений с периферии.
Патологическая боль
Формирование патологической (хронической) боли начинается с усиления потока болевой импульсации с периферии и изменения синаптических процессов, осуществляющих передачу ноцицептивной информации.
Периферическая сенситизация
Вследствие длительного и непрекращающегося тканевого повреждения (воспаления) в тканях происходит накопление алгогенных веществ, среди которых различают (М.Л. Кукушкин, В.К. Решетняк, 1997) тканевые (гистамин, серотонин, простациклины, лейкотриены, цитокины и др.) и плазменные (брадикинин) алгогены, выделяющиеся из эндотелия кровеносных сосудов. В результате происходящих изменений формируется повышенная чувствительность ноцицепторов в месте повреждения, - периферическая сенситизация. Ноцицепторы характеризуются спонтанной активностью, снижением порога возбуждения и повышенной чувствительностью к подпороговым раздражениям (M. Devor, 1996; R. Dubner, 1997).
Клинически периферическая сенситизация выражается в гиперальгезии, т.е. выраженном болевом ощущении при лёгком болевом раздражении и аллодинии, - возникновении болевого ощущения при неболевых (например, тактильных) раздражениях.
Нейрогенное воспаление
При продолжительной болевой стимуляции нормальные сенсорные нейроны становятся гипервозбудимыми, в них появляются участки анормальной активности. Это может происходить в нейронах заднекорешкового ганглия (M. Devor, 1996) и нейронах задних рогов спинного мозга. Возникающие деполяризационные процессы ведут к появлению антидромной, т.е. направленной от спинного мозга к ноцицепторам, стимуляции, нарушению аксоплазматического транспорта трофических веществ (фактор роста нерва), выделению в нервных окончаниях нейрогенных алгогенов (нейропептидов): субстанции Р и кальцитонин-ген-родственного пептида, которые значительно усиливают и поддерживают периферическую болевую стимуляцию. Субстанция Р вызывает скопление и дегрануляцию тучных клеток с высвобождением серотонина и гистамина, и, совместно с кальцитонин-ген-родственным пептидом, воздействует на эндотелий кровеносных сосудов, высвобождая плазменные алгогены: кинины, окись азота и др. (нейрогенное воспаление) (D. Borsook, 1994).
При усилении периферической сенситизации вследствие нейрогенного воспаления боль может появляться при стимуляции хемо- и механосенситивных центростремительных нервов, присутствующих во всех синовиальных и сухожильных тканях, во всех висцеральных органах. В результате этого могут возникать сильные болевые ощущения даже при минимальных движениях и деформациях, что характерно для больных с хроническими дегенеративными заболеваниями костно-мышечной системы, синдромом раздражённой толстой кишки, интерстициальным циститом и другими заболеваниями, сопровождающимися хроническим болевым синдромом.
Повышенная активность ноцицепторов в условиях периферической сенситизации и нейрогенного воспаления ведёт к усилению потока импульсов в центральную нервную систему, что неизбежно ведёт к функциональным изменениям в спинном и головном мозге и возникновению центральной сенситизации.
Центральная сенситизация
В основе центральной сенситизации лежит усиление активации NMDA рецепторов. Установлено, что основным нейротрансмиттером, используемым в болепроводящих системах задних рогов спинного мозга является аминокислота глутамат, молекула которого может связываться с несколькими различными классами рецепторов. На постсинаптической мембране чувствительных нейронов существует два типа глутаматных рецепторов. В восприятие острой боли наиболее вовлечены AMPA (alpha-amino-3-hydroxy-5-methyl-isoxazole-4-propionic-acid)-рецепторы. В случае физиологической боли взаимодействие глутамата с АМРА-рецепторами ведёт к деполяризации мембран нейронов задних рогов спинного мозга и формированию при превышении порога возбуждения потенциала действия.
При длительной, повторяющейся болевой стимуляции, обеспечивающей постоянное и значительное присутствие глутамата в пресинаптическом пространстве, в постсинаптической мембране происходит активация другого типа рецепторов, NMDA (N-methyl-D-aspartate)-рецепторов, что происходит путём вытеснения ионов магния, закрывающих их натриевые и кальциевые каналы (Г.Н.Крыжановский, 1997; М.Л. Кукушкин, В.К. Решетняк, 1997; D. Borsook, 1994).
Активность NMDA-рецепторных комплексов ведёт к возникновению следующих феноменов:
- феномену "взвинчивания" (wind-up phenomen), при котором происходит временная и пространственная суммация возбуждения, что обеспечивает передачу сигнала боли при значительно меньшем количестве глутамата;
- опиоидной толерантности (устойчивости к действию как эндогенных, так и экзогенных опиатов);
- усиленному поступлению ионов натрия и кальция в афферентные терминали и тело нейрона.
Под воздействием ионов кальция активируется фермент NO-синтаза и происходит образование окиси азота (NO), короткоживущего газа, имеющего свободно-радикальные свойства. Окись азота, диффундируя обратно в пресинаптическое пространство, проникает в пресинаптическую мембрану ноцицептора и вызывает, во-первых, опосредованное ингибирование её калиевых каналов, тем самым вызывая торпидность к действию эндогенных опиатов, и, во-вторых, стимулирует высвобождение субстанции Р, биологически активного химического соединения, выполняющего функцию нейротрансмиттера.
Субстанция Р связывается со специфическими NK-1 (нейрокинин-1)-рецепторами, активация которых ведёт к повышению концентрации ионов кальция внутри клетки, повышению возбудимости постсинаптической мембраны и её стойкой деполяризации. Активность нейрокинин-1 рецепторов стимулирует регенерацию нерва и, как показано на моделях животных, продукцию c-fos онкогенного протеина, признанного в настоящее время маркёром патологической боли. C-fos протеин облегчает проведение болевых импульсов и его распространение в высшие отделы центральной нервной системы (таламус), способствует значительному расширению болевого ощущения за пределы дерматома первоначального периферического поражения (D.Brookoff, 1999).
Повышенное выделение глутамата в пресинаптическое пространство, длительная активность NMDA-рецепторов, образование окиси азота и c-fos онкогенного протеина, другие нейрохимические процессы способствуют гиперактивации, истощению и гибели нейронов, формируя очаги (локусы) демиелинизации и дегенерации в различных отделах центральной нервной системы.
Активность системы NMDA-рецепторов и морфофункциональные изменения в периферических нейронах и центральной нервной системе, которые значительно ускоряются и усиливаются при их непосредственном поражении (нейрогенных болях), ведут к повышению возбудимости центральных ноцицептивных структур мозга - центральной сенситизации.
Клинически это выражается в усилении первичной гиперальгезии и аллодинии, появлении зон вторичной гиперальгезии, распространяющихся гораздо шире зон первичного поражения, развитием холодовой и механической гипералгезии, обусловленной развитием феномена "прорастания" (sprouting) терминалей А-волокон в задних рогах спинного мозга (А.В. Новиков, Н.Н. Яхно, 2001).
Возникают локусы (очаги) анормальной электрической активности, обладающие свойством длительно продуцировать поток усиленной импульсации (эктопические анормальные пейсмекеры) (Г.Н. Крыжановский, 1997; М.Л. Кукушкин, В.К. Решетняк, 1997). Это могут быть либо непосредственно невромы, образующиеся при травматическом повреждении нерва, либо другие очаги демиелинизации, регенерации или гибели нервных клеток дорзальных рогов спинного мозга, возникающие при длительной болевой стимуляции.
При последующем развитии патологического процесса такие же очаги образуются не только на уровне спинного мозга, но и в таламических ядрах, соматосенсорной коре больших полушарий головного мозга. Их главное свойство заключается в возможности спонтанной активности и функциональной связи с областью первичного поражения. При их стимуляции у больных вызываются болевые ощущения, по характеру и локализации напоминающие их собственную боль (Г.Н. Крыжановский, 1997).
Формирование генераторов патологически усиленного возбуждения (ГПУВ) и патологической алгической системы (ПАС)
В условиях длительной болевой импульсации возникает слабость и дезинтеграция антиноцицептивных структур мозга, что клинически выражается в низкой эффективности лечения таких
8-09-2015, 23:16