АО «Медицинский университет Астана»
Кафедра информатики и мтематики с курсом медбиофизики
Реферат
“Биофизика мышечного сокращения.”
Выполнила: студентка группы ОМ 129
Берентаева К.
Факультет: Общая медицина
Проверил:Ахметов Б.Г.
Оценка:
Астана 2009
Введение
Физика и биология, на первый взгляд, довольно далекие друг от друга науки.
Но это только на первый взгляд. В действительности же в этих науках есть много общих точек. Например, в анатомии, зрение. Здесь присутствует элемент оптики: лучи света преломляются в хрусталике глаза, и элемент механики: хрусталик деформируется мышцами. Хотя, говоря о мышцах, нельзя не упомянуть о том, что их работа напрямую связана с физикой. Ведь по сути дела, механизм их действия, сокращение в связи с сокращением белковых нитей, физический процесс. А обмен веществ? Ведь питательные вещества переходят из крови в межклеточное вещество, из межклеточного вещества в клетку и из клетки в межклеточное вещество в основном из-за перепада в давлении. А нагревание внешних тканей тела кровью вследствие теплопередачи?
И физика стыкуется с биологией не только в анатомии. У птиц есть аэродинамическое оперение, у рыб гидродинамическая чешуя и боковая линия, для улавливания колебаний воды. Опять же слух…
Интерес биофизики к процессам происходящим в сокращающихся мышцах основан не только на выяснении механизмов мышечных болезней, но и что может быть даже более важным – это раскрытие механизма превращения электрической энергии в механическую, минуя сложные механизмы тяг и передач.
Для того, чтобы понять механизм и биофизические процессы происходящие в сокращающихся мышцах, необходимо заглянуть в строение мышечного волокна.
Структурной единицей мышечного волокна являются Миофибриллы – особым образом организованные пучки белков, располагающиеся вдоль клетки.
Миофибриллы в свою очередь построены из белковых нитей (филаментов) двух типов – толстых и тонких. Основным белком толстых нитей является миозин, а тонких – актин. Миозиновые и актиновые нити – главный компонент всех сократительных систем в организме. Электронно-микроскопическое изучение показало строго упорядоченное расположение миозиновых и актиновых нитей в миофибрилле. Функциональной единицей миофибриллы является саркомер – участок миофибриллы между двумя Z-пластинками. Саркомер включает в себя пучок миозиновых нитей, серединой сцепленных по так называемой М-пластине, и проходящих между ними волокон актиновых нитей, которые в свою очередь прикреплены к Z-пластинам.
1. Молекулярный механизм сокращения
Один грамм ткани скелетной мышцы содержит примерно 100 мг «сократительных белков» – актина и миозина . Механизм их взаимодействия во время элементарного акта мышечного сокращения объясняет теория скользящих нитей, разработанная Хаксли и Хансон.
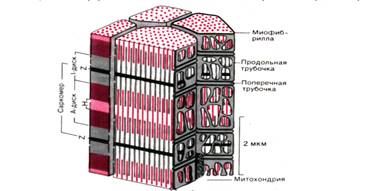
Рис. 1 . Схема участка волокна скелетной мышцы человека (по Garamvolgyi)
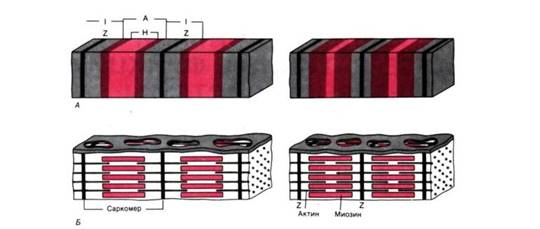
Рис. 2 . А. Поперечнополосатая структура миофибрилл: слева расслабление, справа сокращение. Б. Организация миозиновых и актиновых нитей в расслабленном и сократившемся саркомере. Аддитивный характер укорочения последовательно соединенных саркомеров.
Теория скользящих нитей
Сократительные белки актин и миозин образуют в миофибриллах тонкие и толстые миофиламенты. Они располагаются параллельно друг другу внутри мышечной клетки. Миофибриллы представляют собой сократимые пучки «нитей» (филаментов) диаметром около 1 мкм. Перегородки, называемые Z–пластинками, разделяют их на несколько компартментов–саркомеров длиной примерно по 2,5 мкм.
Согласно теории Хаксли и Хансон поперечная полосатость миофибрилл обусловлена особым взаиморасположением актиновых и миозиновых филаментов. Середину каждого саркомера занимают несколько тысяч «толстых» нитей миозина диаметром примерно по 10 нм. На обоих концах саркомера находятся около 2000 «тонких» (толщиной по 5 нм) нитей актина, прикрепленных к Z–пластинкам наподобие щетинок в щетке. Пучок лежащих в определенном порядке миозиновых нитей длиной 1,6 мкм в середине саркомера выглядит в световом микроскопе темной полосой шириной 1,6 мкм; из–за свойства двойного лучепреломления в поляризованном свете (т. е. анизотропии) она называется А–диском. По обе стороны от А–диска находятся изотропные участки, содержащие только тонкие нити и поэтому выглядящие светлыми; эти так называемые I –диски тянутся до Z–пластинок. Именно в результате такого периодического чередования светлых и темных полос в саркомерах миофибриллы сердечной и скелетной мускулатуры выглядят поперечно–полосатыми.
В покоящейся мышце концы толстых и тонких филаментов обычно лишь слабо перекрываются на границе между А– и I–дисками. Эта зона перекрывания в А–диске выглядит в световом микроскопе гораздо темнее центральной Н–зоны, в которой нет актиновых нитей. На электронных микрофотографиях Н–зоны видна очень тонкая темная М–линия в середине саркомера – сеть опорных белков, по–видимому, удерживающих толстые нити в составе единого пучка.
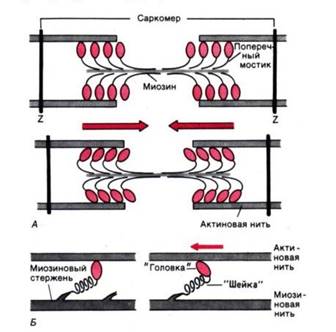
Рис. 3. Функция поперечных мостиков. А. Модель механизма сокращения: миозиновая нить с поперечными мостиками, прикрепленными к соседним актиновым нитям; вверху–до, внизу–после «гребка» мостиков (на самом деле они функционируют асинхронно) . Б. Модель механизма генерирования силы поперечными мостиками; слева– до, справа– после «гребка». Поперечные мостики химически соответствуют субфрагменту миозина–«тяжелому меромиозину», который состоит из субфрагментов I (головка) и II (шейка)
Укорочение саркомеров. Мышца сокращается в результате укорочения множества последовательно соединенных саркомеров в миофибриллах. Сравнивая структуры саркомера в двух различных функциональных состояниях, можно видеть изменения поперечной исчерченности и взаиморасположения нитей во время сокращения: тонкие актиновые филаменты скользят вдоль толстых миозиновых, двигаясь между ними к середине их пучка и саркомера.
Длина нитей не меняется и при растяжении мышцы. Тонкие филаменты попросту вытягиваются из промежутков между толстыми нитями, так что степень перекрывания их пучков уменьшается.
Работа поперечных мостиков . Миозиновые нити несут поперечные, отходящие биполярно, как показано на рис. 3, А, выступы длиной около 20 нм с головками примерно из 150 молекул миозина. Во время сокращения каждая головка (поперечный мостик) может связывать миозиновую нить с соседними актиновыми (рис. 3, А). Движение головок создает объединенное усилие, как бы «гребок», продвигающий актиновые нити к середине саркомера. Сама биполярная организация молекул миозина обеспечивает противоположную направленность (стрелки на рис. 3) скольжения актиновых нитей в левой и правой половинах саркомера.
В результате однократного движения поперечных мостиков вдоль актиновой нити саркомер укорачивается только на 2 х 10 нм, т.е. примерно на 1% своей длины. Однако при изотоническом сокращении мышцы лягушки саркомеры за десятую долю секунды укорачиваются на 0,4 мкм, т. е. на 20% длины. Для этого поперечные мостики должны совершить свои гребковые движения за указанный промежуток времени не один, а 20 раз.
Генерирование мышечной силы. Благодаря упругости поперечных мостиков саркомер может развивать силу даже без скольжения нитей относительно друг друга, т.е. в строго изометрических экспериментальных условиях. Рис.3, Б иллюстрирует такой процесс генерирования изометрической силы. Сначала головка миозиновой молекулы (поперечный мостик) прикрепляется к актиновой нити под прямым углом. Затем она наклоняется под углом примерно 45°, возможно, благодаря притяжению между соседними точками прикрепления на ней и на актиновой нити. При этом головка действует как миниатюрный рычаг, приводя внутреннюю упругую структуру поперечного мостика (видимо, «шейки» между головкой и миозиновой нитью) в напряженное состояние. Возникающее в результате упругое растяжение достигает лишь около 10 нм. Упругое натяжение, создаваемое индивидуальным поперечным мостиком, так слабо, что для развития мышечной силы, равной 1 мН, нужно объединить усилия по крайней мере миллиарда таких соединенных параллельно мостиков.
Даже при изометрическом сокращении поперечные мостики не находятся в непрерывно напряженном состоянии. На самом деле каждая миозиновая головка уже через сотые или десятые доли секунды отделяется от актиновой нити; однако через такое же короткое время следует новое прикрепление к ней. Несмотря на ритмичное чередование прикреплений и отделений с частотой порядка 5–50 Гц, сила, развиваемая мышцей в физиологических условиях, остается неизменной (исключение–летательные мышцы насекомых), так как статистически в каждый момент времени в прикрепленном, обусловливающем напряжение, состоянии находится одно и тоже количество мостиков.
Изометрическое теплообразование. Мышца, поддерживающая определенное сократительное напряжение в изометрических условиях, отличается от сокращающейся изотонически тем, что не выполняет внешней работы (произведение силы на расстояние равно нулю). Однако в каждом цикле прикрепления–отделения поперечных мостиков совершается внутренняя работа по растяжению их упругих структур, которая преобразуется в тепло в момент отсоединения миозиновых головок. Теплота изометрического сокращения («изометрическая работа») за единицу времени возрастает с увеличением количества функционирующих поперечных мостиков и частоты «гребков», требующих расхода АТФ.
Регуляция мышечного сокращения
Обычно мышца возбуждается при поступлении потенциалов действия от иннервирующих мотонейронов; в результате передачи возбуждения через нервно–мышечные синапсы генерируются мышечные потенциалы действия (непрямая стимуляция). Возможна и прямая стимуляция мышечных волокон, но только в экспериментальных условиях. Например, при раздражении изолированной мышцы лягушки одиночным электрическим импульсом длительностью около 1 мс по мышечному волокну от места раздражения примерно через 1–2 мс со скоростью примерно 2 м/с будет распространяться потенциал действия, а еще через несколько миллисекунд оно сократится . Таким образом, сокращение вызывается потенциалом действия, т. е. возбуждением мембраны волокна.
Электромеханическое сопряжение
Передача команды к сокращению от возбужденной клеточной мембраны к миофибриллам в глубине клетки (электромеханическое сопряжение) включает в себя несколько последовательных процессов, ключевую роль в которых играют ионы Са2+ .
Локализация и механизм действия Са2+ . Инъекция Са2+ в мышечные волокна вызывает их сокращение. Интактные живые волокна гораздо меньше подходят для демонстрации прямого воздействия Са2+ на миофибриллы, чем те же волокна после удаления или разрушения поверхностной клеточной мембраны. Для этого их либо «обдирают» («скинируют») механически, либо обрабатывают детергентами, либо используют упоминавшееся выше экстрагирование глицеролом. Такие лишенные сарколеммы («скинированные») мышечные волокна сокращаются только при погружении в раствор, содержащий АТФ и по крайней мере 10–6 М ионизированного кальция для активации АТФазы. В этих условиях поперечные мостики миозиновых нитей могут за счет постоянного расщепления АТФ циклически взаимодействовать с актиновыми нитями. Если активирующий фактор Са2+ удалить из среды (например, добавив связывающие его вещества), миофибриллы расслабляются, поскольку взаимодействие между поперечными мостиками и актином предотвращается, а значит, подавляется активность АТФазы. Такой эффект полностью обратим и в опытах с лишенными сарколеммы волокнами. На ступенчатое повышение концентрации Са2+ от 10– 7 до 10– 5 М они реагируют постепенным увеличением силы сокращения и активности АТФазы, причем оба этих параметра достигают максимума при концентрации Са2+ 10–6 –10–5 М.
Механизм активации ионами кальция мышечного волокна легче понять, рассмотрев структуру актиновых нитей (рис. 4). Каждый такой филамент длиной около 1 мкм и толщиной 5–7 нм состоит из двух закрученных одна вокруг другой цепочек мономеров актина толщиной 5 нм. Похожая структура получится, если взять две нити бус и скрутить их в виде спирали по 14 бусин в каждом витке
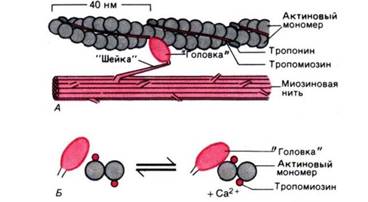
Рис. Действие Cal+ во время активации миофибриллы. А. Актиновая и миозиновая нити на продольном сечении волокна. Б. Они же на его поперечном сечении. Когда Са2+ связывается с тропонином, тропомиозин попадает в желобок между двумя мономерами актина, обнажая участки прикрепления поперечных мостиков.
Через регулярные промежутки примерно по 40 нм актиновые цепочки несут сферические молекулы тропонина, а в желобках между двумя цепочками лежат нити тропомиозина. Исследования с помощью рентгеноструктурного анализа (малоугловое рентгеновское рассеяние) показали, что в отсутствие Са2+ , т.е. при расслабленном состоянии миофибрилл, длинные молекулы тропомиозина располагаются так, что блокируют прикрепление поперечных миозиновых мостиков к актиновым нитям. И напротив, под влиянием Са2+ молекулы тропомиозина глубже опускаются в желобки между цепочками мономеров актина, открывая участки прикрепления для поперечных мостиков. В результате те прикрепляются к актиновым нитям (рис. 4, Б), расщепляется АТФ и развивается мышечная сила.
Такой механизм активации обусловлен действием Са2+ на тропонин, который работает как «кальциевый переключатель»: при связывании с Са2+ его молекула деформируется таким образом, что как бы заталкивает тропомиозин в желобок между двумя цепочками актиновых мономеров, т. е. в «активированное положение».
Хранение и высвобождение ионов кальция . Расслабленная мышца содержит более 1 мкмоль Са2+ на 1 г сырой массы. Если бы соли кальция не были изолированы в особых внутриклеточных хранилищах, обогащенные его ионами мышечные волокна находились бы в состоянии непрерывного сокращения.
Структура внутриклеточных систем хранения кальция в разных мышцах не вполне одинакова (скелетная мышца человека (рис. 1.; мышца лягушки–рис. 5). Во многих участках поверхностная мембрана мышечной клетки образует углубления в виде трубочек (диаметром 50 нм), перпендикулярных продольной оси волокна; эта система поперечных трубочек соединяется с внеклеточной средой и обычно окружает каждую миофибриллу на уровне Z–пластинок (у лягушки) или в области I–дисков (у высших позвоночных).
Перпендикулярно поперечным трубочкам, т. е. параллельно миофибриллам, расположена система продольных трубочек (истинный саркоплазматический ретикулум). Пузырьки на их концах (терминальные цистерны) прилегают к мембранам системы поперечных трубочек, образуя так называемые триады. В этих пузырьках и хранится внутриклеточный кальций. В отличие от поперечной системы продольная не сообщается с внеклеточной средой. Мембраны саркоплазматического ретикулума содержат работающий на энергии АТФ кальциевый насос, который осуществляет активный транспорт из миоплазмы в продольные трубочки, снижая таким образом примерно до 10–7 М миоплазматическую концентрацию этих ионов в покоящейся (расслабленной) мышце.
Электромеханическое сопряжение происходит посредством распространения потенциала действия по мембранам поперечной системы внутрь клетки. При этом возбуждение быстро проникает в глубь волокна, переходит на продольную систему и в конечном счете вызывает высвобождение Са2+ из терминальных цистерн во внутриклеточную жидкость, окружающую миофибриллы, что и ведет к сокращению
При одиночном импульсе сокращение кратковременно расслабление мышцы вызывается обратным переносом активирующих ионов Са2+ посредством кальциевого насоса в каналы саркоплазматического ретикулума. Удаление ионов Ca2+ из миоплазмы идет до тех пор, пока их концентрация в ней не упадет до примерно 10–7 М. При этом подавляются активность АТФазы миозина и взаимодействие между актином и поперечными мостиками, которые отделяются от актиновых нитей.
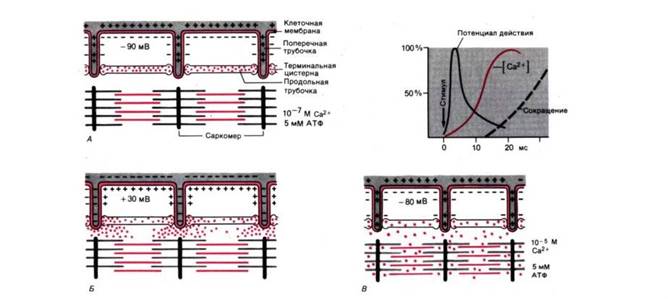
Рис. 5. Схема электромеханического сопряжения. А. Расслабленное мышечное волокно с поляризованной клеточной мембраной. Концентрация Ca2+ в нем ниже 10– 7 М. Б. Потенциал действия меняет полярность мембраны клетки и поперечных трубочек на противоположную; Ca2+ начинает выходить из терминальных цистерн. В. К моменту исчезновения потенциала действия внутриклеточная концентрация Ca2+ достигала примерно 10– 5 М, и саркомеры миофибрилл укоротились. Справа вверху: временная последовательность событий при электромеханическом сопряжении от «латентного» периода до начала сокращения (портняжная мышца лягушки при 0°С)
Распространение возбуждения вглубь волокна.
Этот процесс составляет первый этап электромеханического сопряжения (рис. 6). Воздействуя через микроэлектрод слабыми импульсами тока на мышечное волокно лягушки, эти авторы вызывали локальную деполяризацию такого маленького участка плазматической мембраны, что стимулировалась только одна поперечная трубочка (на уровне Z–пластинки). Возникающее в результате местное сокращение (контрактура) ограничивалось саркомерами поверхностных миофибрилл, непосредственно прилегающих к этой трубочке.По мере усиления стимула активировались все глубже расположенные миофибриллы. Очевидно, мембраны поперечных трубочек легко возбуждаются электрическим током, способны проводить возбуждение и составляют важное звено в процессе передачи сигнала от клеточной мембраны к хранилищам кальция.
Только за счет такой электрической передачи по поперечной системе возможна быстрая мобилизация запасов кальция в глубине волокна, и только этим можно объяснить очень короткий латентный период между стимулом и сокращением. Диффузия Ca2+ от поверхностной мембраны к миофибриллам, находящимся в центре мышечного волокна толщиной 100 мкм, продолжалась бы гораздо дольше, так что для волокон скелетных мышц подобный механизм можно исключить уже по временным соображениям.
Высвобождение кальция при одиночном сокращении. Блинке с коллегами выделили из светящихся медуз белок экворин, который при взаимодействии с Ca2+ излучает свет. После инъекции этого белка изолированное мышечное волокно закрепляли изометрически и раздражали электрическим током с интервалами 100 или 200 мс. С помощью высокочувствительного фотометра (фотоумножителя) регистрировалась люминесценция (излучение света) экворина, сопровождавшая внутриклеточное высвобождение Ca2+ (рис. 7). При стимуляции с частотой 5 Гц она была кратковременной, поскольку ионный насос вскоре перекачивал высвобожденный в миоплазму Ca2+ обратно в саркоплазматический ретикулум; при таком режиме мышца совершает одиночные сокращения. Однако при ритмичном раздражении с частотой 10 Гц (второй стимул поступает уже через 100 мс после первого) волокно расслабляется не полностью. Второе сокращение накладывается на остаточное сокращение после первого стимула, третье – на предыдущие и т. д.
Суммация одиночных сокращений ведет к росту как максимального напряжения в сократительном цикле, так и остаточной величины одиночных сокращений, хотя внутриклеточный уровень Ca2+ после каждого из них (судя по люминесценции) почти возвращается к уровню покоя.
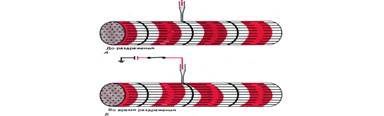
Рис. 6. Опыт, демонстрирующий возможность локальной активации Т–системы. Слабое локальное раздражение микрокатодом волокна поперечнополосатой мышцы лягушки (в области Z–пластинки, непосредственно над Т–трубочкой) вызывает укорочение прилегающих I–дисков: А– до, Б– во время раздражения
При этом опыт, представленный на рис. 7, показывает, что увеличение общего напряжения при стимуляции с интервалами по 100 мс нельзя объяснить повышением уровня внутриклеточного Ca2+ .
Высвобождение Са2+
при тетанусе
.
Если стимулы поступают с высокой частотой (не менее 20 Гц), уровень Са2+
в интервалах между ними остается высоким, потому что кальциевый насос не успевает вернуть все ионы в продольную систему саркоплазматического ретикулума. Как показывает рис. 7, в таких условиях отдельные сокращения почти полностью сливаются. Это состояние устойчивого сокращения, или тетанус, наблюдается в том случае, когда промежутки между стимулами (или потенциалами действия в клеточной мембране) меньше примерно 1/3 длительности каждого из одиночных сокращений. Следовательно, частота стимуляции, необходимая для их слияния, тем ниже,
8-09-2015, 23:43