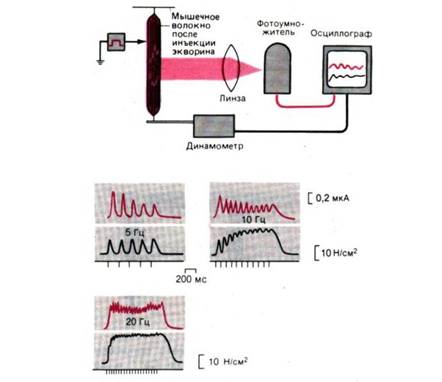
Рис. 7. Опыт, демонстрирующий внутриклеточное высвобождение Са2+ в мышечных волокнах. Люминесценция (красные кривые) и развитие изометрического напряжения в изолированном мышечном волокне шпорцевой лягушки при прямом раздражении импульсами тока длительностью по 0,5 мс с частотой 5, 10 и 20 Гц (моменты раздражения показаны штрихами под кривыми). Заметны суммация и слияние одиночных сокращений до (зубчатого) тетануса при повышении частоты стимуляции. Изометрическое напряжение измерено в ньютонах на 1 см2 площади поперечного сечения мышцы, а люминесценция, вызванная Ca2+ , в единицах силы анодного тока фотоумножителя. Вверху: схема экспериментальной установки, использованной Блинксом и др.
Закон «все или ничего». Этому закону подчиняются быстрые волокна скелетных мышц.
Подпороговые стимулы не вызывают потенциалов действия и высвобождения Ca2+ . Как только интенсивность стимула превысит определенный пороговый уровень, генерируется распространяющийся потенциал действия и происходит максимальное высвобождение Ca2+ , это обеспечивает максимальную силу сокращения, уже не возрастающую при повышении интенсивности стимула.
Вместе с тем при электрическом раздражении целой мышцы сила ее сокращения зависит от интенсивности стимула. Например, если он едва превышает пороговый уровень, ответ по типу «все или ничего» наблюдается только в волокнах, находящихся вблизи от электрода, где плотность тока максимальна; для возбуждения всех волокон требуется гораздо более сильный (максимальный) стимул. Таким образом, только сверхмаксимальное раздражение может равномерно и достаточно надежно активировать изолированную целую мышцу.
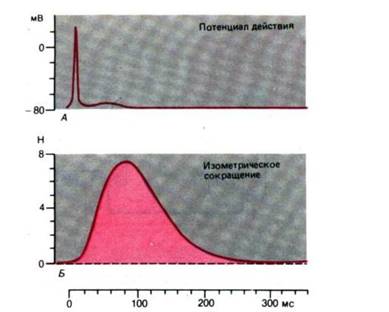
Рис. 8. Временной ход потенциала действия и изометрического сокращения поперечнополосатой мышцы (приводящей большой палец)
Закон «все или ничего» не означает, что ответ раздражаемого мышечного волокна будет всегда одинаков по величине. Например, если мышца только что расслабилась после тетануса, одиночный стимул часто вызывает гораздо более сильное одиночное сокращение, чем до такого «кондиционирования». Причины этой посттетанической потенциации известны так же мало, как и механизм мышечною утомления–снижения силы сокращения при ритмической стимуляции. В обоих случаях у потенциалов действия нормальная амплитуда. При кислородной недостаточности и в еще большей степени при нарушении метаболизма иодацетатом ритмическая стимуляция сопровождается не только снижением силы сокращения, но и замедлением расслабления; в конечном итоге, когда запас АТФ истощается, такая «отравленная» мышца вообще утрачивает способность к расслаблению—становится ригидной. Состояние необратимой ригидности и тетанус следует отличать от различных видов длительного напряжения, обсуждаемых ниже.
Литература:
1. « Физиология человека» в 3т. /под ред. Шмидт Р., Тевс Г., Москва: «Мир», 2004, 326 с.
2. Биофизика, книга 1. Учебное пособие для вузов. /под ред. А.Б.Рубина. - Москва: "Высшая школа", 1987., 365 с.
План
1. Введение
2. Молекулярный механизм сокращения
3. Теория скользящих нитей
4. Укорочение саркомеров.
5. Работа поперечных мостиков
6. Генерирование мышечной силы.
7. Изометрическое теплообразование.
8. Регуляция мышечного сокращения
9. Электромеханическое сопряжениеЛокализация и механизм действия Са2+
10.Распространение возбуждения вглубь волокна.Закон «все или ничего».
8-09-2015, 23:43