Виявилося також, що досвід Гальвані зі скороченням без металів не вдається, якщо м'яз отпрепарован дуже ретельно, без ушкоджень. Але якщо м'яз попередньо поранити, то накидання на неї нерва веде незмінно до скорочення, якщо нерв торкається як пораненої, так і неушкодженої частини.
Найбільша різниця потенціалів, що доходить на м'язі до 0,08 V, спостерігається слідом за нанесенням ушкодження. Протягом першої години струм ушкодження падає майже до половини своєї вихідної величини. У нерві виникаюча при ушкодженні різниця потенціалів звичайно не перевищує, 0,03 V і з часом убуває швидше, ніж у м'язі. У нерві людини вона складає лише близько 0,006 V, що зв'язано, мабуть, з достатком з'єднанних прошарків, що є побічним замиканням для струму, що відводиться від нерва. У процесі дегенерації нерва величина виникаючого при ушкодженні струму перетерплює поступове зменшення, і через 10 днів у теплокровних чи через 30 днів у холоднокровних тварин він уже не виявляється.
Перший прямий доказ переривчастої, коливальної природи порушення в нерві дане Н. Е. Введенським (1883), що примінив телефон для вислуховування струмів дії нерва. Відведення двох крапок нерва на телефон дозволило йому вислухувати ритми діяльності нерва при роздратуваннях різної сили і частоти. Користаючись цією методикою, Н. Е. Введенський ще в 80-х роках минулого століття зробив ряд основних висновків про ритми порушення; ці висновки надалі були підтверджені іншими дослідниками в умовах графічної реєстрації.
Для графічної реєстрації струмів дії, що звичайно представляються у виді швидких електричних коливань, пізніше були застосовані спеціальні гальванометри чи електрометри, що володіють дуже малою інерцією системи, що реєструє. Такими є капілярний електрометр, струнний гальванометр і осцилографи.
Капілярний електрометр у своїй істотній частині представляє капілярну скляну трубку, що містить ртуть зі слабким розчином сірчаної, кислоти що знаходиться, над нею. На поверхні зіткнення ртуті з кислотою виникає різниця потенціалів, причому ртуть заряджається позитивно, а кислота негативно. Наявність заряду зменшує поверхневий натяг ртутного меніска унаслідок взаємного відштовхування однойменно заряджених часток ртуті. Якщо через цю систему пропустити струм, наприклад, у такому напрямку, щоб меніск з'явився катодом, а сірчана кислота — анодом, то заряд меніска зменшиться. Завдяки цьому, поверхневий натяг ртуті збільшиться і меніск опуститься, змістившись в напрямку струму, що пропускається. При зворотному напрямку струму збільшення заряду ртутного меніска зменшить його поверхневий натяг і меніск підніметься. Таким чином, зсув ртутного меніска відбуваються завжди в напрямку діючого струму. Рух ртутного меніска можна спостерігати за допомогою мікроскопа чи фотографувати на світлочутливий папір.
Інерція ртутного меніска усе-таки відносно велика, і більш досконалим приладом є струнний гальванометр. Дуже тонка провідна нитка цього гальванометра, натягнута вертикально між полюсами електромагніта, може відтворювати без значних перекручувань до 500 коливань у секунду.
Значно більшу частоту коливань можуть відтворювати без перекручування шлейфні, чи магніто-електричні, осцилографи, що знайшли собі електрофізіологічне застосування. Рухливою системою тут є шлейф - петля з металевої стрічки, перекинута від одного затиску до іншого через блок, що відтягається догори пружиною. Петля міститься між полюсами постійного магніту. При проходженні струму обидві половини петлі відхиляються в протилежні сторони, тому що напрямок струму в них по-різному. Унаслідок цього вся петля повертається на деякий кут, величина якого визначається силою підводимого струму. Відповідно зміщається пучок світла, відбиваний укріпленим на петлі дзеркальцем. Відхилення світлового пучка фотографуються. Такі осцилографи можуть відтворювати без перекручування до 10 000 коливань у секунду.
Однак безпосереднє використання шлейфних і катодних осцилографів для реєстрації струмів дії є неможливим унаслідок недостатньої чутливості цих приладів. Тому струми дії, що підводяться до осцилографу, необхідно значно підсилювати. Це досягається за допомогою підсилювачів.
Струм дії порозумівається, як уже було зазначено, тим, що збуджена ділянка тканини виявляється на короткий час електронегативною стосовно
спочиваючих ділянок. Коли один з електродів, що
відводять, прикладений до неушкодженої, а інший - до ушкодженої
ділянки нерва чи м'яза, різниця потенціалів між електродами при проходженні хвилі порушення під першим електродом на короткий час
зменшиться: прилад, що реєструє, знайде коливання, що швидко протікає в напрямку, зворотному напрямку струму спокою. Що ж стосується ушкодженої ділянки (під другим електродом), то хвиля збудження, що приходить вгасає в ньому, не змінюючи його електричного стану. У цьому випадку струм дії має форму однофазного, тобто однобічного коливання (рис.4, а).
Якщо ж обидва електроди розташувати на неушкодженій подовжній поверхні препарату, то струм дії представиться у виді двофазного коливання (рис.4, б), тому що ділянки тканини, на яких лежать електроди, з'єднані з гальванометром, не одночасно охоплюються порушенням. Поширюючись з визначеною швидкістю, збудження спочатку доходить до найближчого електрода і створює тут швидко минаючу електронегативність; прилад, що реєструє, дає коливання в одному напрямку. Коли ж при подальшому своєму поширенні порушення, минавши область першого електрода, досягає ділянки, що лежить під другим електродом, у зовнішньому ланцюзі виникає короткочасний струм зворотного напрямку, що регіструється у виді другого коливання, спрямованого в протилежну сторону.
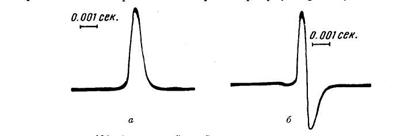 |
Рис.4. Ліворуч однофазний, праворуч двофазний струм дії сідничного нерва жаби, зареєстрований на катодному омосцилографі.
Тривалість однофазного струму дії в кістякових м'язах хребетних тварин виражається тисячними частками секунди: у м'язах жаби вона складає 0,003-0,004 секунди, у м'язах теплокровних - 0,002-0,003 секунди. У нервах тривалість струму дії складає 0,0005-0,0020 секунди. Ніж скоріше проводиться порушення в нерві, тим швидше протікає струм дії.
4. Походження струмів спокою і струмів дії
Ще Сєченов відзначав значення обміну речовин у виникненні різниці потенціалів у тваринних тканинах. У розвиток цих поглядів В. Ю. Чаговец (1896) показав можливість пояснення електричних явищ у м'язах і нервах, виходячи з представлень про концентраційні струми, обумовлених різною концентрацією деяких іонів (наприклад, Н-іонів) у різних ділянках тканини. Різниця концентрацій може бути результатом зміни обмінних процесів у ділянках збудження чи ушкодження.
Пізніше одержала поширення мембранна теорія (Ю. Бернштейн, 1902), заснована на схематичних представленнях про існування на поверхні м'язових і нервових волокон особливого прикордонного шару (мембрани), що, подібно деяким штучно одержуваним перетинкам, є прохідною для катіонів і непрохідною для аніонів. Тому що концентрація електролітів по обох сторонах мембрани неоднакова, то катіони (наприклад, що містяться у відносному надлишку усередині волокна іони калію) прагнуть перейти в зовнішнє середовище, але утримуються на зовнішній поверхні мембрани зарядами своїх аніонів. Унаслідок цього мембрана стає поляризованою: зовнішня поверхня її заряджена позитивно, а внутрішня – негативно( рис 5.). Поки волокно не ушкоджене, по всій його поверхні потенціал однаковий, але при його ушкодженні, тобто при порушенні цілості мембрани, негативні іони одержують можливість дифундувати зсередини назовні: ушкоджене місце виявляється електронегативним стосовно нормально поляризованої поверхні.
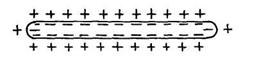
Рис.5. Схема неушкодженого поляризованого нервового волокна (у стані спокою).
Таке походження струму спокою. Збудження ж зв'язане зі збільшенням проникності мембран, про що свідчить ряд спостережень. Завдяки цьому оболонка в ділянці порушення стає однаково проникною для всіх іонів і деполяризується, унаслідок чого збуджена ділянка виявляється електронегативною стосовно іншій позитивно зарядженій поверхні волокна. На відміну від деполяризації, що створюється ушкодженням, деполяризація, зв'язана з порушенням, є оборотною і поширюється разом зі збудженням. Саме поширення збудження з цього погляду представляється, як поширення плани деполяризації. На границі між деполяризованою і поляризованою ділянкоювиникає внаслідок різниці потенціалів переміщення іонів, що приводить до деполяризації нової ділянки і до відновлення поляризації колишнього (рис.6).
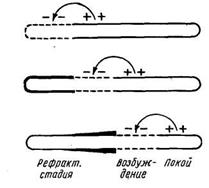
Рис.6. Схема, що показує деполяризацію мембрани нервового волокна в ділянці збудження (ця ділянка показана переривчастою лінією); збудження поширюється ліворуч праворуч і залишає за собою рефракторну фазу (охоплена нею ділянка обкреслена жирною лінією). Стрілки показують виникнення місцевого струму.
8-09-2015, 20:08