Однако не вся вкусовая сенсорная информация передается в корковые центры. Часть ее по выходе из ядра солитарного пучка передается, по видимому, через ретикулярную формацию на пищевой, дыхательной и сосудодвигательный центры, в частности, на ядра блуждающего и языкоглоточного нервов, и обуславливает такие акты, как рвоту, глотание, изменение частоты дыхания и сердцебиений.
Что касается саморегуляции вкусовой сенсорной системы, Ио она несомненно имеется, но механизмы ее почти не выяснены. Вообще, белых пятен в структурно-функциональной организации вкусовой сенсорной системы немало и потому объяснение многих сторон ее деятельности строится на предположениях. Достоверно установлено, что острота вкусовых восприятий зависит от состояния рецепторов желудка. Доказано, что число активно функционирующих вкусовых луковиц у людей в голодном состоянии значительно больше, чем в сытом. Эти данные свидетельствуют о наличии центробежных механизмов регуляции вкусовой сенсорной системы, в том числе регуляции ее периферической части. Однако, как передаются эти влияния на периферическую часть, пока не ясно, так как в составе барабанной струны не обнаружено эфферентных нервных волокон, передающих импульсы на рецепторные вкусовые клетки, а на самих рецепторных клетках не обнаружены эфферентные синапсы. Высказываются предположения, что эфферентные влияния на вкусовые рецепторы осуществляются через симпатическую нервную систему, но передаются непосредственно на вкусовые клетки, а на нервное сплетение под основаниями вкусовых луковиц.
В периферической части обонятельной сенсорной системы имеет место латеральное торможение – при возбуждении одной рецепторной клетки часть других клеток тормозится. Это обусловлено тем, что одно чувствительное волокно, разветвляясь на многочисленные терминали, вступает в синаптические связи с большим числом рецепторных клеток. При возбуждении одной рецепторной клетки в данном нервном окончании возникают нервные импульсы, которые проводятся антидромно по всем разветвлениям этого волокна и блокируют образование импульсов во всех других терминалях. Механизмы латерального торможения более обстоятельно будут рассмотрены при изложении кожной части соматосенсорной системы.
Механизмы центрального торможения во вкусовой сенсорной системе практически не выяснены. В литературе указывается наличие центробежных эфферентных влияний из коры на клетки аркуатного ядра зрительного бугра. Что касается эфферентных влияний на нейроны ядра солитарного пучка, а тем более на чувствительные вкусовые нейроны (в частности узла коленца), то они пока не обнаружены.
Наконец, имеются данные, что некоторые пищевые вещества, например глюкоза, всосавшись в кровь в ротовой полости, буквально за несколько секунд достигают вкусовых центров и оказывают на их клетки определенное влияние, а исходящие из этих центров эфферентные импульсы влияют на промежуточные центры, а возможно и на периферическую часть (через симпатическую нервную систему), изменяя порог чувствительности вкусовых клеток и их адаптации к данному раздражителю.
В заключение следует отметить. Что ощущение вкуса вызывается не только действием четырех основных вкусовых стимулов на вкусовые клетки, но также действием на рецепторы прикосновения, температуры, давления, запаха. Следовательно, целостное ощущение вкуса является результатом совместного функционирования нескольких сенсорных систем вкусовой, экстероцептивной, интероцептивной и обонятельной.
3. Трансдукция вкусового сигнала
Основополагающей функцией вкусовых рецепторных клеток является распознавание вкусовых веществ (ионов и молекул, содержащихся в пище) - трансдукция - и кодирование информации об их концентрации и вкусовой модальности для дальнейшего анализа в соответствующих структурах мозга (Gilbertson et al., 2000; Herness and Chen, 2000). Для трансдукции вкусовых стимулов в настоящий момент общепризнанна множественность механизмов, которые можно разбить на две условные группы. Во-первых, предполагается, что при воздействии солёных и кислых стимулов для образования и передачи сигнала во вкусовой клетке достаточно модуляции ионных потоков через апикальную мембрану, обусловленной изменением ионного состава среды (Na+, в случае солёного; Н*, в случае кислого стимула) во вкусовой поре (Lindemann, 1996; Herness and Gilberson, 1999). Во-вторых, большинство сладких и горьких веществ (а так же ряд аминокислот) активируют хеморецепторный путь передачи сигнала. Предполагается, что в этом случае внешний стимул связываясь с трансмембранным G-белок связывающимся рецептором (GPCR, G-protein coupled receptor) (Lindemann, 2001), переводит его в возбужденное состояние, находясь в котором рецептор катализирует диссоциацию нескольких сотен гетеротримерных G-белков на а-субъединицу и комплекс Ру-субъединиц. Каждый из них может регулировать активность ионных каналов и/или эффекторных ферментов, которые генерируют внутриклеточные сигналы, что в свою очередь приводит к передаче вкусового сигнала далее от вкусовой клетки. (Herness and Gilberson, 1999; Lindemann 2001; Margolskee 2002). Следует отметить, что прогресс последнего десятилетия в области молекулярной биологии, иммуногистохимии и электрофизиологии вкусового органа позволил идентифицировать многие эффекторные молекулы его сигнальных каскадов. Так, одними из первых были идентифицированы восемь а-субъединиц G-белков, которые являются своего рода визитной карточкой сигнального каскада. Наиболее представлено семейство Gi/Go, четыре представителя которого экспрессируются во вкусовых клетках: а-гастдуцин, а-трансдуцин. О гастдуцине: поведенческие эксперименты и регистрации от вкусового нерва показывают, что мыши, у которых была подавлена (knock out) экспрессия гастдуцина, оказались на два порядка менее чувствительны к горьким и сладким (но не к соленым и кислым) веществам по сравнению с мышами дикого типа (Wong et al., 1996). Это однозначно говорит о принципиальной роли гастдуцина в формировании горького и сладкого вкуса. Методом single cell RT-PCR анализировалась экспрессия субъединиц G-белков, и было показано, что гастдуцин, скорее всего, состоит из а-гастдуцина и Ру-субъединиц (Huang et al., 1999), причем а-субъединица, скорее всего, активирует фосфодиэстеразу (PDE), а Ру-комплекс контролирует фосфолипазу С (PLC) (Margolskee, 2002). Гастдуцин экспрессируется преимущественно в клетках типа II (Boughter
et al., 1997; Sbarbati et al., 1999; Smith et al., 1999; Yang et al., 2000b), что является необходимым условием для нормального рецептирования сладких и горьких стимулов (Wong et al., 1995; Ruiz-Avila et al., 2001). Поэтому гасдуцин-положительные клетки типа II могут быть хеморецепторными, однако они не формируют синапсы с эфферентными волокнами; возможно, они секретируют сигнальные молекулы, чтобы, например, модулировать активность соседних вкусовых клеток.
Эти и перечисленные выше данные говорят о том, что потенциально возбуждение вкусовых клеток может происходить путем модуляции активности ионных каналов G-белками или в результате изменения внутриклеточной концентрации циклических нуклеотидов, или за счет генерации инозитолтрифосфата (1Р3) и мобилизации Са2+ (Ogura et al., 2002). Действительно, многие элементы циклонуклеотидного и фосфоинозитидного каскадов найдены во вкусовых клетках, и в частности: аденилатциклаза (AC) (Abaffy, et al., 2003), PDE (Kinnamon and Margolskee, 1996), циклонуклеотид-зависимые каналы (Kaupp and Seifert, 2002, Wei et al., 1998), PLC (Asano-Miyoshia et al., 2000, Rossler et al., 1998), 1Р3-рецепторы (Yan et al., 2001) и TRP-каналы (Zhang, et al., 2003). Однако вклад этих каскадов в возбуждение вкусовых клеток химическими стимулами фактически не исследован и мало известно о том, какие рецепторы контролируют активность этих сигнальных систем.
В 2000г было идентифицировано семейство генов кодирующих так называемые T2R/TRB-penenTopbi (Adler et al, 2000), специфически экспрессирующиеся во вкусовых клетках, и, по-всей видимости, функционирующие как вкусовые рецепторы (Lindemann, 2001). Со структурной точки зрения, T2R/TRB рецепторы лишь отдаленно связаны с другими GPCR, такими как VIR (рецепторы на феромоны), и даже внутри семейства степень гомологии составляет 30-70%. Эти рецепторы имеют высококонсервативные области в цитоплазматических петлях и прилежащих к ним трансмембранных сегментах (предположительно они выполняют функцию взаимодействия с G-белками) и сильно дивергирующие экстраклеточные области (потенциальные сайты связывания с лигандами) (Gilbertson, et al., 2000). У крыс и мышей T2R/TRB рецепторы экспессируются в 15 - 20% вкусовых рецепторных клеток желобоватых и листовидных сосочков и в очень малом количестве вкусовых рецепторных клеток грибовидных сосочков (Margolskee, 2002). Основываясь на данных in situ гибридизации, показано, что T2R/TRB рецепторы экспрессируются в определенных типах вкусовых рецепторных клеток (Adler et al., 2000). В частности, T2R/TRB рецепторы экспрессируются в клетках экспрессирующих гастдуцин (Gilbertson, et al., 2000; Margolskee, 2002). В биохимических экспериментах было показано, что T2R5 - рецептор, распознающий горькое вещество циклогексимид, селективно активирует гастдуцин, но не другие G-белки, экспрессируемые во вкусовых клетках (Chandrachekar et al., 2000). Это дает основание думать, что именно гастдуцин связывает T2R5 и возможно другие рецепторы к горьким веществам с эффекторными ферментами. Было также показано, что горькие вещества стимулируют синтез 1Р3 и что этот ответ блокируется антителами к Ру-субъединицам гастдуцина и к РЬСРг в. то время как контрольные антитела и антитела к а-гастдуцину были неэффективны (Rossler et al., 2000). Это говорит о том, что после рецептор-стимулируемой диссоциации гастдуцина именно ру-комплекс активирует PLCp2, запуская тем самым образование 1Р3 и диацилглицерола (DAG).
Методами биоинформатики в геноме мыши был найден новый рецептор, T1R3, который был идентифицирован как продукт гена из локуса sac на четвертой хромосоме мыши (Max et al., 2001). Поскольку мутации в этом гене ведут к потере чувствительности к сахарину и не затрагивают чувствительность к веществам других вкусовых модальностей, было предположено, что T1R3 - сладко-чувствующий рецептор. Он имеет гомологию порядка 30% с T1R1 и T1R2, GPCR, идентифицированными ранее (Hoon et al., 1999), которые структурно составляют еще одно семейство вкусовых рецепторов (T1R1, T1R2 и Т1Ю). Интересно отметить, что методом двойной in situ гибридизации было показано, что T1R1 и T1R2 эксперессируются в разных вкусовых клетках, но некоторые вкусовые клетки экспрессируют комбинации T1R1 и T1R3 или T1R2 и T1R3 (Hoon et al., 1999; Nelson et al, 2001). Ha основании этих данных было сделано предположение, что функциональный рецептор, скорее всего, является гетеродимером (T1R1/T1R3 и T1R2/T1R3), что нашло подтверждение в следующих экспериментах. Оказалось, что при экспрессии комбинации T1R2 + T1R3 в клетках НЕК-293 (которые в контроле не реагировали на вкусовые вещества) таковые отвечали на сладкие вещества (Nelson et al., 2001). При экспрессии комбинации T1R1 + T1R3 индуцировалась чувствительность к аминокислотам (Nelson et al., 2001). Это дает основание полагать, что гетеродимер T1R2/T1R3 является сладко-чувствующим рецептором, а T1R2/T1R3 - функционирует как рецептор аминокислот.
Вкус умами вызывается L-глутаматом в миллимолярных концентрациях (Lindemann, 2001). Во многих тканях глутамат идентифицирован как возбуждающий нейротрансмиттер, где в микромолярных концентрациях он активирует ионотропные и метаботропные (т.е. G-белок сопряжённые) рецепторы (Brown and Dale, 2002). Классические глутаматные рецепторы также найдены во вкусовых клетках (Chaudhari et al., 1996; Lin and Kinnamon, 1999), но они вряд ли обеспечивают рецепцию глутамата как вкусового вещества, поскольку афинность этих рецепторов к глутамату характеризуется микромолярными константами диссоциации. Недавно был идентифицирован подтип метаботропного рецептора (mGlu4t (truncated)), специфически экспрессируемый во вкусовых клетках (Chaudhari et al., 2000). Его особенность в том, что по сравнению с классическим mGlu4-рецептором у него не хватает 130 аминокислот на N-конце (расположен экстраклеточно и несет сайт связывания глутамата), что сдвигает афинность mGlu4t рецептора в область миллимолярных концентраций (Chaudhari et al., 2000). Это обстоятельство, а также апикальная локализация mGlu4t, дают основание считать, что данная изоформа метаботропного рецептора, специфическая для вкусовых клеток, обеспечивает детекцию глутамата в пище (Chaudhari et al., 2000).
Итак, во вкусовых клетках найдены многие элементы внутриклеточных сигнальных систем - рецепторы, G-белки, эффекторные ферменты, ионные каналы. На этом основании до недавнего времени доминировала гипотеза, предполагающая одновременное существование множественных механизмов, использующих cGMP/cAMP и IP3/DAG/Ca в качестве вторичных мессенджеров для трансдукции вкусовыми клетками стимулов разных вкусовых модальностей (Lindemann, 1996; Herness and Gilberson, 1999; Gilberson et al., 2000; Lindemann 2001; Margolskee 2002). Здесь следует отметить сложность изучения молекулярных механизмов вкуса, которая состоит в том, что многочисленные модели вкусовой трансдукции трудно проверить экспериментально, поскольку в электрофизиологических экспериментах не более 5% вкусовых клеток отвечает на адекватные вкусовые стимулы. Однако недавно появившиеся данные ставят под сомнение эту традиционную точку зрения. Согласно поведенческим экспериментам и регистрации от вкусового нерва, у животных с нокаутированными PLC ((32 изоформой) или ионным каналом TRPM5 (семейства TRP-каналов; Runnels et al., 2002, Vazquez et al., 2001, Venkatachalam et al., 2001), практически исчезала чувствительность к сладким, горьким стимулам и аминокислотам при неизменной чувствительности к кислым и солёным стимулам (Zhang et al., 2003). Следовательно, PLCP2 и TRPM5 являются общими и ключевыми элементами сигнальных каскадов, обеспечивающих трансдукцию сладких, горьких стимулов и аминокислот.
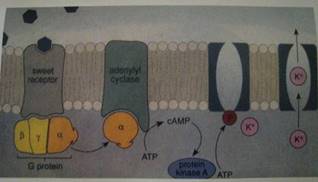
Рис.1 Трансдукция веществ, обладающих сладким вкусом
4. Вкусовые нарушения
вкусовой анализатор трансдукция окраска
4.1 Общие принципы
Анатомическое разнообразие системы вкуса, а также присутствие разнообразных резервных механизмов способствует значительной защите системы от полной и постоянной потери вкусового восприятия. Например, потеря нескольких периферических вкусовых очагов не вызовет изменений в способности ротовой полости в целом воспринимать вкус (Mott, Grushka and Sessle 1993). Система восприятия вкуса в гораздо большей степени восприимчива к вкусовым искажениям или восприятию фантомного вкуса. Например, дисгевзия является заболеванием намного более распространенным в условиях профессиональных воздействий, чем потеря вкусовых ощущений. Хотя способность восприятия вкуса рассматривается в качестве более устойчивой к возрастным изменениям, чем обоняние, специалистами были описаны случаи ослабления вкусовых ощущений с возрастом. При раздражении слизистой рта может иметь место временная потеря вкуса. Теоретически, это может быть вызвано воспалением вкусовых клеток, закупоркой вкусовых пор или функциональными изменениями на поверхности вкусовых клеток. Ожог может повлиять на кровоснабжение языка и таким образом повлиять на систему восприятия вкуса. Система распространения слюны также может быть повреждена. Раздражители могут вызвать отек, что приведет к закрытию слюнных канальцев. Поглощенные слюнными железами токсичные вещества во время выделения могут нанести вред тканям канальцев. Любой из этих процессов может вызвать продолжительную сухость во рту, в результате чего будет повреждена система восприятия вкуса. Воздействие токсичных веществ может повлиять на скорость обновления вкусовых клеток либо внести изменения в их внутреннюю или внешнюю химическую среду. Известно множество веществ, рассматривающихся в качестве нейротоксинов и способных нанести ущерб непосредственно периферийным вкусовым нервам или повредить вышележащие вкусовые анализаторы мозга.
4.2 Повреждающие агенты
Пестициды Пестициды применяются довольно широко и их остатки можно обнаружить в мясе, овощах, молоке, дождевой и питьевой воде. И хотя наибольшему риску подвергаются рабочие, вовлеченные в процессы производства и использования пестицидов, остальное население также подвергается их воздействию. Основные пестициды включают хлорорганические, фосфорорганические соединения и соединения солей карбаминовой кислоты. Хлорорганические соединения чрезвычайно устойчивы и поэтому сохраняются в окружающей среде в течение долгих периодов времени. Прямая интоксикация данными соединениями вызывает у людей поражение нейронов центральной нервной системы. Вследствие меньшей устойчивости в окружающей среде фосфорорганические пестициды получили более широкое применение, однако, они также являются и более токсичными, чем хлорорганические соединения; ингибирование ацетилхолинэстеразы может вызвать отклонения неврологического и поведенческого характера. Токсичность пестицидов солей карбаминовой кислоты примерно равна описанной токсичности для фосфорорганических соединений, и они часто используются там, где применение последних не приводит к ожидаемым результатам.Воздействие пестицидов ассоциировалось с устойчивым горьким или металлическим вкусом (Schiffman and Nagle 1992), дисгевзией неопределенного характера (Ciesielski et al. 1994) и реже с потерей вкуса. Пестициды могут попадать к вкусовым рецепторам через воздух, воду и пищу, а также могут проникать через кожу, пищеварительный тракт, конъюнктиву и дыхательные пути. Поскольку большое количество пестицидов растворимы в липидах, они могут легко проникать через липидные оболочки организма. Поражение системы восприятия вкуса может носить опосредованный характер, не зависящий от места исходного воздействия; в эксперименте на мышах после инъекции пестицидов в кровяной русло на языке у животных были выявлены данные инсектициды. После воздействия пестицидов были выявлены изменения в морфологии вкусовых сосочков. Были также отмечены дегенеративные изменения в окончаниях сенсорных нервов, которые вполне могут объяснить случаи нарушений нервной передачи, которые были также зарегистрированы. Металлическая дисгевзия может представлять собой форму сенсорной парестезии, вызванную воздействием пестицидов на вкусовые сосочки и расположенные в них центростремительные нервные окончания. Существуют, однако, некоторые доказательства того, что пестициды могут вмешиваться в работу медиаторов и таким образом препятствовать передаче вкусовой информации в центральную нервную систему (El-Etri et al. 1992). У рабочих, которые подвергались воздействию фосфорорганических пестицидов, могут иметь место неврологические нарушения, независимые от снижения содержания холинэстеразы в крови и выявляемые при проведении электроэнцефалографии и нейропсихологического теста. Предполагается, что эти пестициды вне зависимости от воздействия на холинэтестразу подвергают мозг нейротоксическому воздействию. Согласно результатам исследований, с воздействием пестицидов связывают увеличение слюновыделения, но до сих пор остается неясным, какие вкусовые изменения это может повлечь за собой.
Металлы и литейная лихорадка
Изменения в восприятии вкуса наблюдались у пострадавших при воздействии некоторых металлов и их соединений, включая ртуть, медь, селен, теллур, цианистый калий, ванадий, кадмий, хром и сурьму. Металлический привкус также появлялся у рабочих, которые подверглись воздействию паров цинка или окиси меди, этот же привкус наблюдался в случаях отравлений, связанных с приемом внутрь солей меди, либо в случаях воздействий выбросов, образующихся при применении газового
8-09-2015, 20:13