
Рис.2. Радикальные и нерадикальные продукты ПОЛ [Aibert W, Girotty, 1998]
Особенно важно подчеркнуть, что происходит нарушение мембранных структур, изменение транспорта ионов Са2+, создающее угрозу жизни клетки [Michiel, Remacle, 2004].
1.3. Антиоксидантная система .
Образование АФК, известных как прооксиданты, наблюдается во многих метаболических процессах и является обязательным атрибутом нормальной аэробной жизни. Функционирование и развитие клеток, а так же организма в целом, в кислородсодержащем окружении не могло бы быть возможным без существования защитных систем, основу которых составляют ферментативные и неферментативные антиоксиданты [Петрович, Гуткин, 2005]. Постоянное образование прооксидантов в живых организмах уравновешено их дезактивацией антиоксидантами, поэтому для поддержания гомеостаза необходима непрерывная регенерация антиоксидантной способности. Отсутствие или сбои этой непрерывности сопровождаются накоплением окислительных повреждений и приводят к возникновению окислительного стресса [Меньщикова с соавт., 2006].
Основными функциями антиоксидантной системы являются: ограничение интенсивности реакции свободнорадикального и перекисного окисления; защита чувствительных к окислительным повреждениям биомолекул мембран, внутри - и внеклеточных структур от действия свободных радикалов и перекисных соединений; восстановление окислительных молекулярных повреждений [Шепелев с соавт., 2004]. В целом основная задача системы антиоксидантной защиты состоит в предотвращении и ограничении развития патологических состояний, вызываемых окислительными повреждениями структур организма [Владимиров, 1998; Журавлев, 2003].
Общепринятой номенклатуры антиоксидантов в настоящее время нет. По химической природе биоантиокислители представляют собой широкий класс соединений: ферменты (СОД, каталаза, глутатионпероксидаза (ГПО)), фенолы и полифенолы (токоферолы, эвгенол), флавоноиды (рутин, кверцетин), стероидные гормоны и многие другие соединения [Бурлакова, Храпова, 2004]. В зависимости от растворимости различают жирорастворимые (витамин Е, А, К, убихинон) и водорастворимые (витамин С, SH-содержащие соединения) биоантиокислители, по молекулярной массе выделяют группу низкомолекулярных антиоксидантов (глутатион, α-токоферол, мочевая кислота) и высокомолекулярных ( ферритин, каталаза) [Кения, Лукаш, Гуськов, 1993].
К числу энзимных антиоксидантов относят прежде всего супероксидредуктазу (СОР), восстанавливающую О2- в пероксид водорода, СОД, катализирующую реакцию дисмутации О2- с образованием пероксида водорода и молекулярного кислорода [Cord, Fridovich, 2000], каталазу, восстанавливающую Н2О2, глутатионзависимые пероксидазы и трансферазы (ГТ) [Wendel, 2004].
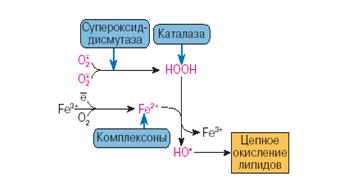
Рис.3. Антиоксиданты водной фазы [Владимиров, 1998]
Главной ферментативной системой плазмы крови является ГПО внеклеточных жидкостей и ГПО гидроперекисей липидов (ГПО 4), которая будучи липофильным соединением эффективно взаимодействует с гидроперекисями фосфотидилхолина, холестерина и эфиров холестерина в липопротеинах низкой плотности (ЛПНП), восстанавливая их, следовательно, защищая от окислительной модификации [Thomas at al., 2006; Abiaka, Al-Awadi, Olusi, 2003]. Кроме того ГПО 4 совместно с токоферолом практически полностью подавляет ПОЛ в биологических мембранах благодаря тому, что витамин Е эффективно восстанавливает пероксирадикалы, а фермент разлагает гидроперекиси, препятствуя тем самым их вовлечению в окислительный цикл [Ursini, Bindoli, 2005; Бондарь, Ланкин, Антоновский, 1989].
ГТ, в отличие от селенсодержащей ГПО, для которой лучшими субстратами являются гидрофильные гидроперекиси с малым размером молекул, эффективно восстанавливает гидрофобные гидроперекиси с большим объемом молекулы: гидроперекиси полиненасыщенных жирных кислот (линолевой и арахидоновой), фосфолипидов [Бобырев, Почерняева, Стародубцев, 2005; Колесниченко, Кулинский, 2004].
Вместе с тем во всех водных и липидных фазах организма могут протекать радикальные окислительные процессы, в защите от которых важную роль играют антиоксиданты-ингибиторы органических радикалов, среди которых важное место занимают соединения фенольного типа. В настоящее время выделено несколько тысяч фенольных соединений, среди которых выраженным антиоксидантным эффектом обладают витамины Е и К, убихиноны, триптофан и фенилаланин, а так же большинство растительных (флавоноиды) и животных пигментов [Дубина, 2005].
В антирадикальной защите липопротеинов плазмы крови и клеточных мембран α-токоферолу принадлежит ведущая роль – одна его молекула защищает ≈10000 молекул ненасыщенных жирных кислот [Евстигнеева, Волков, Чудинова, 2003], при этом считается, что α-токоферол способен обезвредить не менее 60% образующихся пероксильных радикалов [Тиунов, 1995]. Окисление α-токоферола со свободными радикалами компенсируется биорегенерацией молекул этого антиоксиданта в реакциях восстановления коантиоксидантами (АН), редокс-потенциал которых ниже, чем у радикала α-токоферола (α-Тф-О.) [Бурлакова, Крашков, Храпова, 1998]. В результате такой реакции не только происходит восстановление витамина Е, но и предотвращается возможность инициации α-токофероксильным радикалом окисления липидов:
α-Тф-О. + RH→ α-Тф-ОH + R. (1)
α-Тф-О. + АH→ α-Тф-ОH + А. (2)
В физиологических условиях вторая реакция обычно превалирует над первой, так как константа скорости реакции α-Тф-О. с НЖК не превышает 103М-1с-1, в то время как для реакции (1) константа скорости может достигать значений 104-107М-1с-1.К наиболее изученным коантиоксидантам относятся убихинол, аскорбиновая кислота (АК), билирубин [Менщикова, Зенков, 1993].
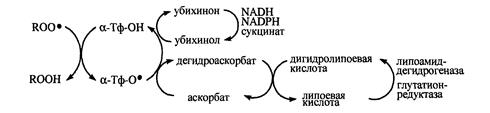
Рис.4. Окислительно-восстановителные превращения α-токоферола и сопряженных с ним коантиоксидантов [Меньщикова с соавт., 2006].
АК превосходит другие антиоксиданты плазмы в защите липидов от перекисного окисления, так как только это соединение достаточно реакционноспособно, чтобы эффективно ингибировать инициацию ПОЛ в водной фазе [Padayatty, Katz, Wang, 2003].
Важными внеклеточными антиокислителями являются альбумины – простые гидрофильные белки плазмы крови, которые являются основными носителями SН групп. SН содержащим соединениям принадлежит ведущая роль в защите клеток от ОН-радикала, образующегося в реакции Фентона или в результате разложения молекул воды под действием ионизирующих излучений.
Необходимо отметить наличие антирадикальных свойств у белков, хелатирующих ионы железа и других металлов с переменой валентностью. В первую очередь это трансферрин, лактоферрин и церулоплазмин. Основная роль трансферрина и близкого ему по структуре лактоферрина, содержащегося в молоке и выделяющегося фагоцитами при их активации, состоит в акцептировании «свободного» железа, что препятствует образованию радикалов НО.в реакциях Фентона, катализируемых ионами Fe [Зборовская, Банникова, 2000].Значимую роль в этом процессе играет церулоплазмин, Cu-содержащий белок, обеспечивающий окисление Fe 2+ до Fe3+ и делающий тем самым железо доступным для связывания трансферрина [Воскресенский с соавт., 2004; Krsek-Staples, Wbster, 2004; Василец, 2004]
Наиважнейшим антиоксидантом внеклеточной жидкости является мочевая кислота (МК). Ввиду высокого ее содержания в плазме крови человека некоторые исследователи считают, что на нее приходится 35-65% защиты липопротеинов от окисления, 10-15% ингибирования НО. и 12% ингибирования синглетного кислорода. Кроме того МК может выступать синергистом с радикалами α-токоферола и аскорбиновой кислотой, что усиливает их антиоксидантное действие [Булгакова, 2006].
Глава 2. Материалы и методы .
2.1 Объект исследования .
Объектом исследования явилась замороженная плазма крови относительно здоровых небеременных и беременных женщин, поставляемая из Хакасского республиканского центра планирования семьи. Плазма хранилась при температуре -20° С до определения концентрации диеновых коньюгатов и малонового диальдегида. Всего было обследовано 127 женщин, деление на группы приведено в таблице 1.
Таблица 1
Группы обследованных женщин
| Сроки беременности | МДА, ДК |
| Контроль | N=31 |
| I триместр | N=29 |
| II триместр | N=34 |
| III триместр | N=33 |
2.2. Определение содержания малонового диальдегида .
В липидных системах в результате процессов ПОЛ образуется МДА, взаимодействие которого с 2-тиобарбитуровой кислотой (ТБК) приводит к образованию хромогена с максимумом поглощения в красной области видимого спектра при длине волны 532 нм. [Стальная, 1997].
Реактивы:
1. 30%-ная трихлоруксусная кислота (ТХУ);
2. 0,1 М этилендиаминтетрауксусная кислота (ЭДТА);
3. 1%-ная тиобарбитуровая кислота;
4. 0,05 н раствор NaOH;
5. Физиологический раствор.
Ход определения:
К 0,2 мл образца добавляли 0,8 мл физиологического раствора и 0,5 мл ТХУ. Перемешивали, оставляли стоять на льду 2 часа. Затем центрифугировали 15 мин. при 3000 об/мин. 1 мл супернатанта переносили в другую пробирку, добавляли 0,075 мл 0,1 М ЭДТА и 1%-ного ТБК, растворенной в 0,05 Н растворе NaOH. Содержимое перемешивали и ставили в кипящую водяную баню на 15 минут. Затем пробирки охлаждали при комнатной температуре и измеряли поглощение при длинах волн 532нм, а затем при 600 нм.
Расчет МДА производят по формуле:
D532 – D600
C= _________________ x f,
E
где:
C – концентрация МДА, мкмоль/л плазмы (эритроцитов),
D532 – оптическая плотность при длине волны 532 нм;
D600 – оптическая плотность при длине волны 600 нм;
E – коэффициент экстинции 1,56x105М-1см-1;
f – коэффициент разведения. (f=9,94)
2.3. Определение содержания диеновых коньюгатов .
Вследствие π-π переходов спектры конъюгированных гидроперекисей полиненасыщенных жирных кислот характеризуются интенсивным поглощением в ультрафиолетовой области спектра с максимумом при 232-234 нм.
Определение содержания ДК проводили в экстрактах эритроцитов. Для этого липиды из эритроцитов экстрагировали стократным избытком смеси растворителей (гептан-изопропанольная смесь в соотношении 1:1). В гомогенизатор вносили 0,1 мл. упакованных эритроцитов, добавляли 5 мл. изопропилового спирта и тщательно растирали до получения гомогенной суспензии. Содержимое гомогенизатора количественно переносили в мерную центрифужную пробирку, в которую затем добавляли 5 мл. гептана.
Экстракт центрифугировали в течение 10 минут при 1700g. Надосадочную фракцию переносили в градуированную пробирку и добавляли 1/5 объема 0,74%-ного водного раствора KCI для отмывки липидного экстракта от нелипидных примесей. После тщательного встряхивания образовавшаяся эмульсия расслаивалась на две прозрачные фазы. В гептановом экстракте (верхняя фаза) измеряли спектрофотометрически содержание сопряженных диенов в кювете с длиной оптического пути 1,0 см против гептана. Расчет количества ДК производили, используя молярный коэффициент экстинкции при длине волны 233 нм, равный 27000 М¯¹*см¯¹ [Паранич и соавт., 1993] и выражали в ммолях на мл клеток.
С=Д/К*F
Д – оптическая плотность;
К – коэффициент ммолярной экстинкции = 21,0 мМ-1 см-1;
F – фактор разведения.
2.4. Статистическая обработка результатов
Статистическую обработку результатов осуществляли общепринятыми методами с использованием пакета прикладных программ Statistica 7.0. Достоверность различий оценивали по непараметрическому критерию Манна-Уитни, с достоверностью Р<0,05 [Лакин, 1980].
Глава 3. Результаты исследований и обсуждение .
Говоря об общей опасности воздействия свободных радикалов на организм, нельзя обойти эту проблему и у беременных женщин, от здоровья которых зависит будущее целого поколении детей.
При неосложненной беременности в организме женщин происходит целый ряд адапционно-приспособительных процессов, направленных на обеспечение адекватного течения гестационного периода, роста и развития плода. Известно, что важными составляющими адаптивных перестроек являются показатели агрегатного состояния крови, иммунного статуса, эндокринной системы а так же перекисного окисления липидов. В их сбалансированном взаимодействии в ответ на регулирующее влияние гипоталамуса заложено качество адаптации организма [Гусак, 2006].
Физиологическая беременность может сопровождаться существенными изменениями в про/антиоксидантном статусе. Одними из параметров, которые позволяют оценить состояние свободнорадикальных процессов является диеновые коньюгаты и малоновый диальдигид. В нормальных условиях активность этих процессов находится на невысоком уровне, но при стрессовых ситуациях происходит усиленная активация процессов ПОЛ под действием АФК, что приводит к патологическому состоянию.
3.1. Содержание продуктов перекисного окисления липидов в плазме крови небеременных женщин .
В данной работе было определено содержание диеновых коньюгатов и малонового диальдегида у небеременных женщин (N=31), которые составили группу контроля. Согласно литературным данным содержание продуктов ПОЛ в частности МДА, являющегося одним из конечных продуктов данного процесса, с возрастом неуклонно возрастает [Банкова, Никанорова, 1988]. В группе небеременных женщин возраст значительно варьировал: от 18 до 39 лет. В связи с этим, мы разделили женщин контрольной группы по возрастному критерию на две подгруппы: в первую подгруппу вошли женщины в возрасте 18-28 лет (N=18), во вторую – 29-39 лет (N=13).
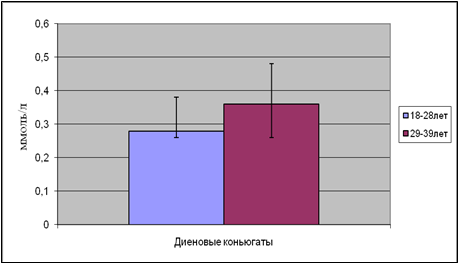 |
Содержание ДК и МДА в этих подгруппах приведены на рисунках 5 и 6.
Рис.5. Содержание диеновых коньюгатов в возрастных подгруппах
В возрастной подгруппе от 18 до 28 лет содержание ДК составило 0,28 ммоль/л, а в возрастной подгруппе от 29 до 39 лет концентрация увеличилась на 28%. Та же закономерность выявлена и в содержании МДА.
Концентрация МДА в первой подгруппе составила 0,85мкмоль/л, во второй подгруппе уровень МДА повысился на 31%.
Наблюдаемые изменения свидетельствуют о том, что с возрастом на фоне усиления процессов ПОЛ происходит угнетение антиоксидантной системы. Нарушения в системе перекисного окисления липидов и антиоксидантной активности являются одним из механизмов формирования антиоксидантной недостаточности вследствие чрезмерного усиления ПОЛ.
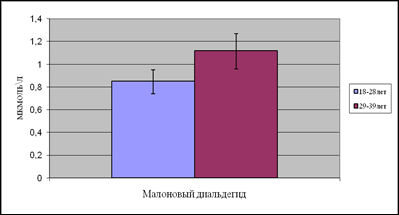
Рис.6. Содержание малонового диальдегида в возрастных подгруппах
В результате активации ПОЛ и накопления свободных радикалов происходит окислительная модификация липопротеинов плазмы крови, нарушение структурно-функциональной целостности клеточных мембран, освобождение лизосомальных ферментов, что в конечном итоге приводит к патологическим процессам в клетке и организме в целом.
При анализе содержания ДК и МДА между двумя возрастными подгруппами достоверных отличий не выявлено. Вследствие чего при сравнительном анализе содержания продуктов ПОЛ у беременных и небеременных женщин будут использованы данные по общей контрольной группе. Результаты исследования содержания ДК и МДА в общей контрольной группе приведены на рисунке 7.
Уровень ДК в контрольной группе женщин составил 0,35 ммоль/л, а МДА – 0,96 мкмоль/л. Наши данные по содержанию МДА в плазме крови согласуются с приведенными в литературе [Гусак, 2006]. Данных по содержанию ДК в плазме крови у относительно здоровых женщин мы в доступной литературе не встретили.
Анализ полученных результатов показал, что уровень ДК, являющихся первичными продуктами ПОЛ, существенно превышает таковой для конечного продукта – МДА.
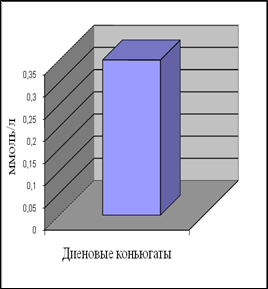
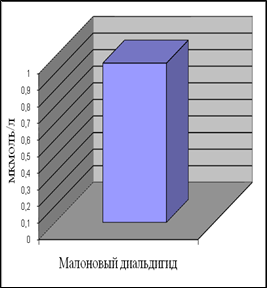
Рис.7. Содержание продуктов перекисного окисления липидов в плазме крови небеременных женщин
Согласно данным литературы, у женщин показатели перекисного окисления липидов, а так же антиоксидантной системы изменяются в динамике репродуктивного цикла (1, 7, 14, 21 дни) [Гусак, 2006]. В первые дни менструального цикла происходит увеличение соотношения адреналин/серотонин, что свидетельствует о преобладании в функциональной активности гипоталамуса адренергических компонентов. Это приводит к депрессии противосвертывающей и антиокислительной систем крови, а так же активации клеточного и гуморального иммунитета, повышению неспецифической иммунной резистентности. Что проявляется увеличением концентрации диеновых коньюгатов и малонового диальдегида, активацией свертывающего потенциала крови.
К 21 дню цикла активность АОС прогрессивно увеличивается, на фоне этого снижается количество продуктов ПОЛ, в том числе ДК и МДА. Это необходимо для обеспечения процессов имплантации в слизистую матки, развития плаценты и создания оптимальных условий, которые позволят, с одной стороны – обеспечить надежную защиту организма женщины, а с другой – вынашивание беременности. Данная закономерность имеет глубокий биологический смысл и направлена на обеспечение условий для реализации репродуктивной функции человека.
В данной работе мы не учитывали возможность влияния динамики репродуктивного цикла женщин на уровень продуктов ПОЛ, хотя это явление могло отразиться на полученных результатах.
3.2. Содержание диеновых коньюгатов и малонового диальдегида и в плазме крови женщин в разные периоды беременности .
Во время гестации, начиная с момента зачатия и до завершения родов, в организме матери возникают интегративные процессы, которые необходимы для поддержания функционального единства организма матери и плода. Эти процессы позволяют выполнить главную задачу, то есть сохранение плода, и заключается в адаптации в I триместре беременности.
Несмотря на то, что в антигенном отношении мать и плод всегда несовместимы, в большинстве случаев после имплантации бластоцисты беременность развивается нормально и завершается родами в срок. Если бы взаимоотношения между матерью и плодом строились по варианту реципиент – аллотрансплантат, то беременность вряд ли продолжалась дольше срока, чем выживание обычного трансплантата. Следовательно, в системе "мать – плод" существуют механизмы, направленные на сопереживание двух антагонистически настроенных субъектов.
В данной работе было определено содержание ДК и МДА в плазме крови у беременных женщин (N=96), разделенных по триместрам, в возрасте от 20 до 28 лет.
Группу сравнения составили небеременные женщины. Результаты определения уровня ДК и МДА приведены на рисунках 8 и 9.
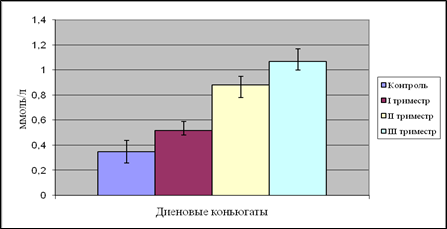 |
Рис. 8. Содержание диеновых коньюгатов в плазме крови женщин в динамике беременности
У женщин в I триместре беременности наблюдается достоверное повышение уровня продуктов ПОЛ по сравнению с контролем. Так, содержание ДК и МДА повысилось на 74 и 77% соответственно. Это можно объяснить тем, что сразу же после зачатия происходит значительная перестройка жизнедеятельности организма беременных женщин, которая сопряжена с изменениями в системах крови, гемостаза, эндокринной, иммунной системах и с изменением биохимического состояния организма в целом. «Не ожидая» подобных изменений, ткани и органы испытывают определенный стресс, в результате которого резко повышается количество свободных радикалов, атакующих, помимо прочего, клетки плаценты и эмбриона.
Увеличение количества свободных радикалов (СР) во время беременности связано с различными причинами. Одной из причин является их участие в синтезе прогестерона. СР активируют процесс перекисного окисления липидов, в результате которого образуются гидроперекиси холестерина, являющиеся предшественниками данного гормона.
Прогестерон является очень важным гормоном, необходимым для поддержания всего течения беременности. Он подготавливает эндометрий матки к имплантации оплодотворенной яйцеклетки, а затем способствует сохранению беременности: подавляет активность гладкой мускулатуры матки, поддерживает в центральной нервной системе доминанту беременности; стимулирует развитие концевых секреторных отделов молочных желез и рост матки, синтез стероидных гормонов; оказывает иммунодепрессивное действие, подавляя реакцию отторжения плодного яйца [Савченко с соавт., 2006].

Рис 9. Концентрация прогестерона при разных сроках беременности
Кроме того, прогестерон является предшественником стероидных гормонов плода, а так же эстрогенов, андрогенов, альдостерона и других гормонов коры надпочечников. Содержание прогестерона в крови беременной женщины увеличивается, повышаясь в 2 раза к 7-8 неделе, а затем более плавно возрастает к 34 недели (Рис 9). Следовательно, интенсификация процессов перекисного окисления липидов во время беременности является физиологически необходимым процессом.
Гормональные связи «пронизывают» все компоненты функциональной системы мать-плацента-плод. Так, в I триместре беременности происходит тесное взаимодействие материнско-плацентарной эндокринной системы, а во II и особенно в III триместрах плод и плацента выступают как общий орган синтеза эстрогенов.
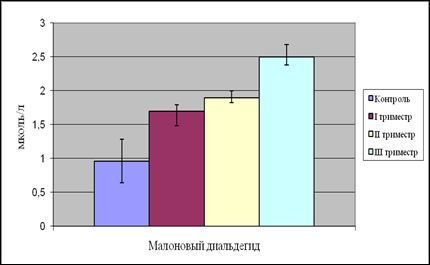
Рис. 9. Содержание малонового диальдегида в плазме крови женщин в динамике беременности
Дальнейший анализ полученных нами данных показал, что уровень как МДА, так и ДК
8-09-2015, 23:05