7. Значение в дыхании импульсов с рецепторов легких, дыхательных мышц, дыхательных путей и скелетной мускулатуры.
Регуляция дыхательного ритма осуществляется нервной системой и гуморальным путем. Центр дыхания располагается в продолговатом мозгу. В дыхательном центре различают два отдела: отдел вдоха и отдел выдоха, функции которых взаимосвязаны. При возбуждении отдела вдоха происходит торможение отдела выдоха и наоборот. Участвуют в регуляции дыхания специальные скопления нервных клеток в мосту и в промежуточном мозге. Кроме того, в спинном мозге находятся группы клеток, отростки которых идут в составе спинномозговых нервов к дыхательным мышцам. В дыхательном центре попеременно возбуждение сменяется торможением. При вдохе легкие расширяются, их стенки растягиваются, что раздражает окончания блуждающего нерва. Возбуждение передается к дыхательному центру и тормозит его деятельность. Мышцы перестают получать возбуждение от дыхательного центра и расслабляются: грудная клетка опускается, ее объем уменьшается и происходит выдох. При расслаблении центростремительные волокна блуждающего нерва перестают возбуждаться (нет тормозящего действия на дыхательный центр), и дыхательный центр, не получая тормозящих импульсов, вновь возбуждается - наступает очередной вдох. Таким образом, происходит как бы саморегуляция: вдох вызывает выдох, а выдох - вдох. Другой причиной изменения деятельности дыхательного центра является концентрация углекислоты в крови. Она является специфическим возбудителем дыхания: повышение концентрации углекислоты в крови (особенно в крови, омывающей дыхательный центр) приводит к возбуждению дыхательного центра - дыхание становится частым и глубоким. Глубокое и частое дыхание продолжается до тех пор, пока концентрация углекислоты в крови не снизится до нормального уровня. На понижение концентрации углекислоты в крови дыхательный центр отвечает понижением возбудимости вплоть до полного прекращения своей деятельности на некоторое время - до установления нормального уровня углекислоты. Ведущим физиологическим механизмом, влияющим на дыхательный центр, является рефлекторный, за которым следует гуморальный. Дыхание подчинено коре головного мозга, о чем свидетельствует факт произвольной задержки дыхания (хотя и на очень короткое время) или изменения частоты и глубины дыхания. Это подтверждается учащением дыхания человека при эмоциональных состояниях. Возбуждение дыхательного центра также может вызвать пониженное содержание кислорода в крови и некоторые лекарственные препараты, поступающие в кровь. С дыханием связаны и такие защитные акты, как кашель и чиханье. Они осуществляются рефлекторно, а их центры находятся в продолговатом мозге. Кашель возникает в ответ на раздражение слизистой оболочки гортани, глотки или бронхов (при попадании пыли, частиц пищи и др.). При кашле после глубокого вдоха воздух с силой выталкивается из дыхательных путей, приводя в движение голосовые связки (возникает характерный звук); вместе с выдыхаемым воздухом удаляются и раздражители. Акт чиханья возникает в ответ на раздражение слизистой оболочки носа; причины его те же, что и при кашле.
Остановка дыхания, которая наблюдается в результате понижения концентрации углекислоты в крови, называется апноэ. Нарушение ритма дыхания - одышка и учащенное дыхание - происходит вследствие повышения концентрации углекислоты в крови и называется диспноэ (гиперпноэ). Возникновением апноэ объясняется способность некоторых людей нырять и оставаться несколько минут под водой. Ныряльщик перед погружением в воду производит несколько частых и глубоких дыхательных движений, вследствие чего в крови понижается концентрация углекислоты. Ныряльщикам за жемчугом удается пробыть под водой до 5 минут, но только после длительных тренировок. А при задержке дыхания на 1-1,5 минуты наступает диспноэ - усиленное дыхание. При задержке дыхания концентрация углекислоты в крови повышается и как следствие - увеличивается возбудимость дыхательного центра до тех пор, пока содержание углекислоты в крови не понизится. С повышенной концентрацией углекислоты в крови связан и первый вдох новорожденного.
8. Особенности процесса саморегуляции дыхания .
Координированные сокращения дыхательных мыщц обеспечиваются ритмической активностью нейронов дыхательного центра. Такие нейроны сгруппированы в целом ряде структур мозгового ствола. Неотъемлемым звеном аппарата регуляции дыхания являются также хеморецепторные и механорецепторные системы, обеспечивающие нормальную работу центрального дыхательного механизма в соответствии с потребностями организма в обмене газов.
Центральный дыхательный механизм мозгового ствола и его афферентные системы обеспечивают поддержание легочной вентиляции, адекватной уровню газообмена в тканях организма, т.е. выполняют гомеостатическую функцию. Дыхательные мышцы участвуют в локомоторных актах, поддержании позы, фонаторной функции. Поэтому нейрональные структуры спинного и продолговатого мозга, управляющие респираторной мускулатурой, находятся под контролем вышележащих центров супрапонтинного уровня – мозжечка, среднего и промежуточного мозга, больших полушарий
Важное значение имеет влияние этих центров в регуляции дыхания, связанной с двигательной активностью организма. Особую роль играет кортикальная регуляция дыхания у человека. Это связано прежде всего с участием дыхательного аппарата в речевой функции. Человек может управлять дыханием произвольно. Это позволяет ему менять частоту и глубину дыхания и даже задерживать его. Но длительность произвольного апноэ ограничена: как только парциальное давление накапливающегося в крови СО2 достигает определенного уровня, сильное возбуждение хеморецепторов (императивнй стимул) выводит дыхание из-под произвольного контроля и дыхательные движения возобновляются.
Центральный дыхательный механизм жестко ограничивает вмешательство в регуляцию дыхания со стороны супрапонтинных структур, охраняя жизненно важную гомеостатическую, газообменную функцию дыхательной системы.
9. Роль в регуляции дыхания коры больших полушарий. Примеры изменения дыхания под влиянием корковой регуляции.
Дыхательный центр в продолговатом мозге находится под влиянием нервных импульсов из опоясывающей извилины, полосатых тел, зрительных бугров, подбугровой области, среднего мозга, мозжечка и премоторных областей больших полушарий головного мозга.
Раздражение лобных долей больших полушарий головного мозга вызывает изменения дыхательных движений (В.Я.Данилевский, 1874). У животных были выработаны условные рефлексы на дыхание. После удаления участков коры больших полушарий, осуществляющих высшую регуляцию дыхания, безусловно-рефлекторные изменения дыхания сохранялись, а затем восстанавливались.
Влияние больших полушарий головного мозга на дыхание доказывается тем, что дыхательные движения могут быть по желанию изменены: дыхание можно замедлить или задержать на некоторое время или, наоборот, участить или углубить. Психические влияния и внушения вызывают изменение дыхательных движений. У людей вентиляция легких увеличивается при действии сигналов, предшествующих мышечной деятельности.
Высшая, наиболее тонкая форма регуляции дыхания – обеспечение соответствия дыхательных движений условиям жизни – осуществляется при участии больших полушарий головного мозга.
Предполагается, что нейроны больших полушарий более возбудимы к повышению напряжения углекислоты в крови, чем дыхательный центр в продолговатом мозге.
10. Регуляция дыхания при мышечной работе, при пониженном атмосферном давлении.
Регуляция дыхания при мышечной работе
При усиленной мышечной работе в крови, протекающей через мышцы, увеличивается давление СО2 , повышается концентрация водородных ионов и температура крови, что благоприятствует отдаче О2 в мышцах.
Эти изменения осуществляются рефлекторно и нервно-гуморально. В результате значительного усиления обмена веществ, поступления в кровь углекислоты, молочной и других кислот и накопления в крови водородных ионов и возбуждения дыхательного центра легочная вентиляция при мышечной работе увеличивается как за счет учащения дыхания, так и за счет углубления.
Парциальное давление СО2 в артериальной крови во время мышечной работы не изменяется или незначительно увеличивается (не соответственно величине физической нагрузки). При тяжелой мышечной работе оно меньше, чем при легкой работе. Возбудимость дыхательного центра к действию СО2 во время мышечной работы не увеличивается. Эти факты доказывают, что ведущая роль в увеличении дыхания при мышечной работе принадлежит не гуморальному, а рефлекторному механизму (рефлексам из химических рецепторов кровеносных сосудов, из механических рецепторов дыхательных мышц, диафрагмы и двигательного аппарата). Особенно возрастает число афферентных импульсов из рецепторов дыхательных мышц и диафрагмы при дыхании в противогазе, что приводит к рефлекторному усилению вентиляции легких. Ведущая роль в изменениях дыхания при мышечной работе принадлежит корригирующим влияниям больших полушарий головного мозга и условным рефлексам, главным образом при действии словесных раздражителей. Условные рефлексы на дыхательный аппарат тем больше выражены, чем ближе начало выполнения трудовых или физических упражнений (предрабочее и предстартовое состояние). Изменения дыхания во время речи обусловлены только нервными влияниями.
Рефлекторная регуляция дыхания в соответствии с интенсивностью мышечной работы связана с изменениями физико-химических свойств крови, влияющих на возбудимость дыхательного центра. Максимальный уровень вентиляции легких ограничен пределом возбудимости дыхательного центра. Повышение этого предела при чрезмерном увеличении притока к нему афферентных импульсов уменьшает вентиляцию легких. При произвольном усилении дыхания обмен веществ в дыхательных мышцах больше, чем при произвольном.
При интенсивной мышечной работе вследствие увеличения вентиляции легких возрастает парциальное давление кислорода в альвеолярном воздухе и резко снижается содержание кислорода в венозной крови в результате увеличения его потребления в мышцах. Поэтому артериально-венозная разность кислорода в крови достигает 160 см3 и больше.
Легочная вентиляция обычно увеличивается параллельно интенсивности физической работы. Чем напряженнее работа, тем больше воздуха проходит через легкие и тем больше газообмен. При гимнастических упражнениях вентиляция легких увеличивается в 2-7 раз, а при беге на средние дистанции – даже до 20 раз.
При интенсивной длительной мышечной работе, беге, езде на велосипеде, плавании нередко «через» некоторое время наступает очень сильная отдышка, затруднение дыхания и ощущение стеснения в груди, учащаются сердцебиения, повышается кровяное давление («мертвая точка»). Это состояние преодолевается волевым усилием, следовательно, импульсами с больших полушарий, и тогда исчезает ощущение стеснения в груди и удушье, а самочувствие улучшается. Дыхание становится ровным и спокойным, значительно повышается работоспособность («второе дыхание»).
При «мертвой точке» в крови накапливаются углекислота и молочная кислота и повышается концентрация водородных ионов. Это вызывает дальнейшее углубление дыхания, резко усиленную вентиляцию легких, которая сопровождается увеличением выделения СО2 в выдыхаемый воздух и падением концентрации водородных ионов; то же наступает и благодаря увеличенному выделению молочной кислоты при потоотделении во время «второго дыхания».
Усиленная вентиляция легких перед интенсивной мышечной работой предупреждает наступление «мертвой точки».
При подъеме тяжестей, лазании, гребле и других физических упражнениях происходит натуживание – закрытие голосовой щели после вдоха и задержка дыхания при напряжении выдыхательной мускулатуры, что приводит к повышению давления в легких и сужению легочных сосудов.
Регуляция дыхания при пониженном атмосферном давлении.
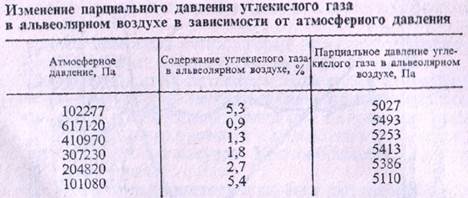 Парциальное давление СО2
в альвеолярном воздухе остается относительно постоянным и при повышении и при понижении атмосферного давления в окружающей среде. Это имеет большое физиологическое значение, так как создает необходимое условие для нормальной возбудимости дыхательного центра и делает возможным дыхание на больших высотах. Постоянство парциального давления СО2
в альвеолярном воздухе обусловлено регулированием дыхания. Это было доказано в опытах, проведенных на людях в стальной камере, сконструированной для изучения влияния высоких и низких атмосферных давлений (барокамера).
Парциальное давление СО2
в альвеолярном воздухе остается относительно постоянным и при повышении и при понижении атмосферного давления в окружающей среде. Это имеет большое физиологическое значение, так как создает необходимое условие для нормальной возбудимости дыхательного центра и делает возможным дыхание на больших высотах. Постоянство парциального давления СО2
в альвеолярном воздухе обусловлено регулированием дыхания. Это было доказано в опытах, проведенных на людях в стальной камере, сконструированной для изучения влияния высоких и низких атмосферных давлений (барокамера).
В отличие от СО2 парциальное давление О2 в альвеолярном воздухе повышается при увеличении атмосферного давления и понижается при уменьшении этого давления.
При подъеме на высоту падает атмосферное давление и уменьшается парциальное давление О2 в альвеолярном воздухе. Это вызывает нарушение снабжения организма О2 . Корличество кислорода в крови становится недостаточным (гипоксемия). Гипоксемия сопровождается значительной потерей углекислоты за счет выдыхания СО2 при увеличении легочной вентиляции.
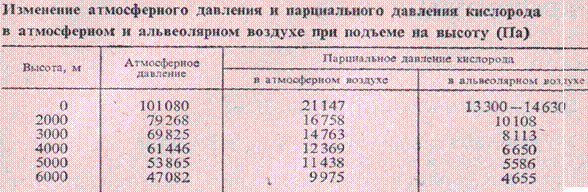
При быстром подъеме на 3-4 км у некоторых здоровых людей начинает развиваться гипоксемия, появляется высотная, или горная, болезнь, выражающаяся в ряде расстройств (головная боль, одышка, усталость, понижение зрения, слуха и др.). У большинства людей горная болезнь отмечается на высоте 4,5-5,5 км. Некоторые люди могут переносить без признаков горной болезни высоту 6 км.
При медленном подъеме на высоту или у живущих на высоте организм успевает приспособиться и не испытывает кислородного голодания (высотная акклиматизация). при этом усиливается легочная вентиляция, что увеличивает парциальное давление О2 в легких и насыщение им артериальной крови. Учащается пульс, увеличиваются минутный объем сердца и кровяное давление, количество эритроцитов и гемоглобина за счет депонированной крови, что повышает кислородную емкость крови на 20-25%. Увеличивается новообразование эритроцитов и содержание в них гемоглобина.
11. Кривая диссоциации-оксигемоглобина. Роль гемоглобина в поддержании газового гомеостаза.
Большая часть О2 и СО2 переносится в форме связи их с гемоглобином в виде молекул HbO2 и HbCO2. Максимальное количество кислорода, связываемое кровью при полном насыщении гемоглобина кислородом, называется кислородной емкостью крови. В норме ее величина колеблется в пределах 16,0—24,0 об% и зависит от содержания в крови гемоглобина, 1 г которого может связать 1,34 мл кислорода (число Хюфнера). В клинике определяют степень насыщения артериальной крови кислородом, представляющую собой выраженное в % отношение содержания кислорода в крови к ее кислородной емкости.
Связывание кислорода гемоглобином является обратимым процессом, зависимым от напряжения кислорода в крови (при понижении напряжения кислорода оксигемоглобин отдает кислород), что отражается так называемой кислородно-диссоциационной кривой гемоглобина, а также от других факторов, в частности от рН крови. Как правило, все эти факторы смещают кривую диссоциации оксигемоглобина, увеличивая или уменьшая ее наклон, но не изменяя при этом ее S-образную форму.
Рис. Кислородно-диссоционная кривая гемоглобина.
![]()
![]()
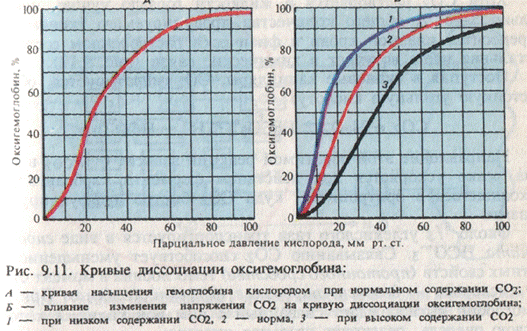
Присоединение О2 к Hb, образование HbO2 и освобождение его (восстановление Hb) зависят от концентрации О2 и в меньшей степени – СО2 . Ни у одного животного напряжение О2 в крови не уравновешено непосредственно парциальным давлением этого газа в атмосфере (т.е. предельным значением О2 в газовой смеси окружающей среды). Дыхательные пигменты у большинства животных насыщаются при значительно более низком значении парциального давления. Диапозон значений парциального давления, в котором осуществляется либо присоединение, либо отщепление О2 является одной из наиболее важных характеристик дыхательного пигмента у различных животных. Эти значения парциального давления определяют условия, в которых функционирует данный дыхательный пигмент.
В области парциальных давлений О2 от 80 до 100 мм рт. ст. (т.е. колебаний, которые могут легко наступить в альвеолярном воздухе в связи с изменением внешней газовой среды организма) кривая диссоциации имеет пологий вид. Это означает, что насыщение гемоглобина кислородом при таких колебаниях парциального давления О2 меняется незначительно (от 93 до 96%). При обычных условиях дыхание Hb почти полностью насыщен О2 . Крутое падение кривой при парциальном давлении О2 , меньшем чем 60 мм рт. ст. (напряжение, характерное для тканей организма), свидетельствует о том, что при этих условиях кровь, протекающая через капилляры, интенсивно освобождает О2 , используемый функционирующими клетками и тканями организма, в которых напряжение О2 невелико. Кривая диссоциации HbO2 имеет S-образную форму. Нижний отрезок кривой определяет свойства Hb в зоне низкого парциального давления О2 , характерного для тканей организма; средняя часть кривой характеризует условия в венозной крови, верхний отрезок – в альвеолах легких.
Диссоциация HbO2 зависят от температуры: при одинаковом парциальном давлении О2 в окружающей среде HbO2, отдает больше О2 при температуре тела организма (37-38ºС), чем при более низкой температуре. Присоединение О2 к Hb в определенной мере регулируется концентрацией СО2 в крови.
СО2 реагируя с водой, образует угольную кислоту Н2СО3, поэтому при повышенном напряжении СО2 кислотность крови увеличивается. Увеличение кислотности крови приводит к уменьшению способности Hb связывать О2 . Такой механизм регуляции количества HbO2, обеспечивает эффективную систему переноса О2 в организме. В капиллярах функционирующих тканей концентрация СО2 достаточно велика, поэтому совместное действие низкого напряжения О2 и высокого напряжения СО2 ведет к освобождению гемоглобином большого количества кислорода. В капиллярах легких (или жабр) напряжение СО2 ниже, и под действием низкого напряжения СО2 и высокого напряжения О2 гемоглобин присоединяет к себе большее количество О2 .
Особенности транспортной функции переносчиков О2
, определяемые различиями в экологических условиях существования различных животных, проявляются в соответствующих изменениях кривой диссоциации HbO2. Кривые диссоциации у амфибий, обитающих главным образом на суше, сдвинуты вправо по сравнению с соответствующими кривыми для видов животных, обитающих в воде. Чем доступнее кислород для животного, тем выше
8-09-2015, 19:51