Содержание
1.Введение
2.Кроветворение
2.1. Эмбриональный гемопоэз
2.2. Постэмбриональный гемопоз
2.3.Регуляция гемопоэза
3. Моноциты
4. Воспаление
4.1. Медиаторы воспаления
4.2. Стадии воспаления
4.2.1. Альтерация
4.2.2. Экссудация
4.2.2.1. Сосудистые реакции и изменение кровообращения
4.2.2.2. Эмиграция лейкоцитов
4.2.2.3. Фагоцитоз
4.2.3. Пролиферация и завершение процесса
5. Монокины
6. Заключение
7. Список литературы
1. Введение
Воспаление является одним из сложнейших процессов часто встречающихся в патологии человека и нередко являющихся причиной многих нарушений жизнедеятельности организма человека и животных.
Воспаление является важной проблемой и предметом изучения всех отраслей медицины и относится к тем феноменам дискуссии о сущности которых на протяжении веков ведут медики, биологи философы. Проблема воспаления стара, как стара и сама медицина.
Однако до сих пор нет единого представления о том, где же место воспаления в биологии, медицине и патологии. Поэтому пока не существует исчерпывающего определения этого процесса.
Но наиболее полным я нахожу именно это определение:
Воспаление – типовой патологический процесс, сформировавшийся в эволюции как защитно-приспособительная реакция организма на воздействие патогенных (флогогенных) факторов, направленная на локализацию, уничтожение и удаление флогогенного агента, а также на устранение последствий его действия и характеризующийся альтерацией, экссудацией и пролиферацией.
В этом реферате я бы хотела посмотреть на воспаление с точки зрения клеточных реакций, в частности, роли лейкоцитов в этом процессе, взаимосвязи между лейкоцитами различных типов.
Как известно, заправилами и регуляторами клеточной миграции, пролиферации, роста сосудов являются моноциты, поэтому, основной акцент в реферате я сделала именно на их роли. Воспаление – типовой патологический процесс, сформировавшийся в эволюции как защитно-приспособительная реакция организма на воздействие патогенных (флогогенных) факторов, направленная на локализацию, уничтожение и удаление флогогенного агента, а также на устранение последствий его действия и характеризующийся альтерацией, экссудацией и пролиферацией.
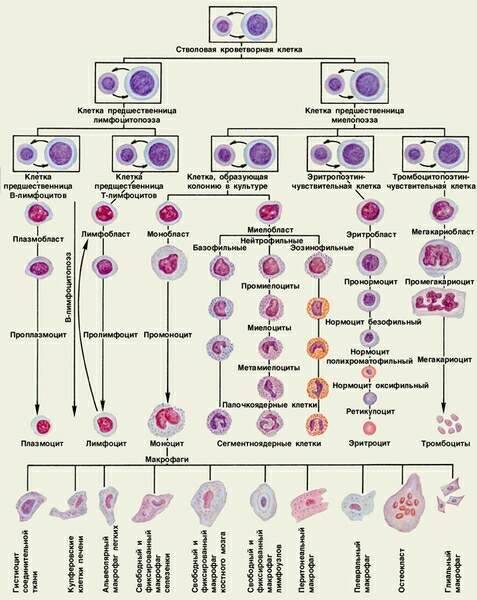
2. Кроветворение
2.1. Эмбриональный гемопоэз
Кроветворением, или гемопоэзом, называют развитие крови. Различают эмбриональный гемопоэз, который происходит в эмбриональный период и приводит к развитию крови как ткани, и постэмбриональный гемопоэз, который представляет собой процесс физиологической регенерации крови.
Развитие эритроцитов называют эритропоэзом, развитие гранулоцитов — гранулоцитопоэзом, тромбоцитов — тромбоцитопоэзом, моноцитов — моноцитопоэзом, развитие лимфоцитов и иммуноцитов — лимфоцито- и иммуноцитопоэзом.
Эмбриональный гемопоэз
В развитии крови как ткани в эмбриональный период можно выделить 3 основных этапа, последовательно сменяющих друг друга – мезобластический, гепатолиенальный и медуллярный.
Первый, мезобластический этап – это появление клеток крови во внезародышевых органах, а именно в мезенхиме стенки желточногомешка, мезенхиме хориона и стебля. При этом появляется первая генерация стволовых клеток крови (СКК). Мезобластический этап протекает с 3-й по 9-ю неделю развития зародыша человека.
Второй, гепатолиенальный этап начинается с 5—6-й недели развития плода, когда печень становится основным органом гемопоэза, в ней образуется вторая генерация стволовых клеток крови. Кроветворение в печени достигает максимума через 5 мес и завершается перед рождением. СКК печени заселяют тимус, селезенку и лимфатические узлы.
Третий, медуллярный (костномозговой) этап — это появление третьей генерации стволовых клеток крови в красном костном мозге, где гемопоэз начинается с 10-й недели и постепенно нарастает к рождению. После рождения костный мозг становится центральным органом гемопоэза.
Рассмотрим подробнее особенности гемопоэза в стенке желточного мешка, в печени, в тимусе, селезенке, лимфатических узлах и в костном мозге.
Кроветворение в стенке желточного мешка
В мезенхиме стенки желточного мешка обособляются зачатки сосудистой крови, или кровяные островки. В них мезенхимные клетки округляются, теряют отростки и преобразуются в стволовые клетки крови. Клетки, ограничивающие кровяные островки, уплощаются, соединяются между собой и образуют эндотелиальную выстилку будущего сосуда. Часть стволовых клеток дифференцируется в первичные клетки крови (бласты). Большинство первичных кровяных клеток митотически делится и превращается в первичные эритробласты, характеризующиеся крупным размером – мегалобласты. Это превращение совершается в связи с накоплением эмбрионального гемоглобина (HbF) в цитоплазме бластов. В некоторых первичных эритробластах ядро подвергается кариорексису и удаляется из клеток, в других ядро сохраняется. В результате образуются безъядерные и ядросодержащие первичные эритроциты, отличающиеся большим размером по сравнению с нормоцитами и поэтому получившие название мегалоцитов. Такой тип кроветворения называется мегалобластическим. Он характерен для эмбрионального периода, но может появляться в постнатальном периоде при некоторых заболеваниях.
Наряду с мегалобластическим в стенке желточного мешка начинается нормобластическое кроветворение, при котором из бластов образуются вторичные эритробласты, из которых образуются вторичные эритроциты (нормоциты).
Развитие эритроцитов в стенке желточного мешка происходит внутри первичных кровеносных сосудов, т.е. интраваскулярно. Одновременно экстраваскулярно из бластов, расположенных вокруг сосудистых стенок, дифференцируется небольшое количество гранулоцитов — нейтрофилов и эозинофилов.
Часть СКК остается в недифференцированном состоянии и разносится током крови по различным органам зародыша, где происходит их дальнейшая дифференцировка в клетки крови или соединительной ткани. После редукции желточного мешка основным кроветворным органом временно становится печень.
Кроветворение в печени
Печень закладывается примерно на 3—4-й неделе эмбриональной жизни, а с 5-й недели она становится центром кроветворения. Кроветворение в печени происходит экстраваскулярно, - по ходу капилляров, врастающих вместе с мезенхимой внутрь печеночных долек. Источником кроветворения в печени являются стволовые клетки крови, из которых образуются бласты, дифференцирующиеся во вторичные эритроциты.
Одновременно с развитием эритроцитов в печени образуются зернистые лейкоциты, главным образом нейтрофильные и эозинофильные.
Кроме гранулоцитов, в печени формируются гигантские клетки — мегакариоциты, - предшественники тромбоцитов. К концу внутриутробного периода кроветворение в печени прекращается.
Кроветворение в тимусе
Тимус закладывается в конце 1-го месяца внутриутробного развития, и на 7—8-й неделе его эпителий начинает заселяться стволовыми клетками крови, которые дифференцируются в лимфоциты тимуса. Увеличивающееся число лимфоцитов тимуса дает начало T-лимфоцитам, заселяющим T-зоны периферических органов иммунопоэза.
Кроветворение в селезенке
Закладка селезенки также происходит в конце 1-го месяца эмбриогенеза. Из вселяющихся сюда стволовых клеток происходит экстраваскулярное образование всех видов форменных элементов крови, т.е. селезенка в эмбриональном периоде представляет собой универсальный кроветворный орган. Образование эритроцитов и гранулоцитов в селезенке достигает максимума на 5-м месяце эмбриогенеза. После этого в ней начинает преобладать лимфоцитопоэз.
Кроветворение в лимфатических узлах
Первые закладки лимфоузлов человека появляются на 7—8-й неделе эмбрионального развития. Большинство лимфатических узлов развивается на 9—10-й неделе. В этот же период начинается проникновение в лимфатические узлы стволовых клеток крови, из которых на ранних стадиях дифференцируются эритроциты, гранулоциты и мегакариоциты. Однако формирование этих элементов быстро подавляется образованием лимфоцитов, составляющих основную часть лимфатических узлов.
Появление единичных лимфоцитов происходит уже в течение 8—15-й недели развития, однако массовое «заселение» лимфатических узлов предшественниками T- и B-лимфоцитов начинается с 16-й недели, когда формируются посткапиллярные венулы, через стенку которых осуществляется процесс миграции клеток. Из клеток-предшественников дифференцируются сначала лимфобласты (или большие лимфоциты), а далее средние и малые лимфоциты. Дифференцировка T- и B-лимфоцитов происходит, соответственно, в T- и B-зависимых зонах лимфатических узлов.
Кроветворение в костном мозге
Закладка костного мозга осуществляется на 2-м месяце эмбрионального развития. Первые гемопоэтические элементы появляются на 12-й неделе развития; в это время основную массу их составляют эритробласты и предшественники гранулоцитов. Из СКК в костном мозге формируются все форменные элементы крови, развитие которых происходит экстраваскулярно. Часть СКК сохраняется в костном мозге в недифференцированном состоянии. Они могут расселяться по другим органам и тканям и являться источником развития клеток крови и соединительной ткани.
Таким образом, костный мозг становится центральным органом, осуществляющим универсальный гемопоэз, и остается им в течение постнатальной жизни. Он обеспечивает стволовыми кроветворными клетками тимус и другие гемопоэтические органы.
2.2. Постэмбриональный гемопоэз
Постэмбриональный гемопоэз представляет собой процесс физиологической регенерации крови, который компенсирует физиологическое разрушение дифференцированных клеток. Он подразделяется на миелопоэз и лимфопоэз.
Миелопоэз происходит в миелоидной ткани, расположенной в эпифизах трубчатых и полостях многих губчатых костей. Здесь развиваются эритроциты, гранулоциты, моноциты, тромбоциты, а также предшественники лимфоцитов. В миелоидной ткани находятся стволовые клетки крови и соединительной ткани. Предшественники лимфоцитов постепенно мигрируют и заселяют тимус, селезенку, лимфоузлы и некоторые другие органы.
Лимфопоэз происходит в лимфоидной ткани, которая имеет несколько разновидностей, представленных в тимусе, селезенке, лимфоузлах. Она выполняет функции образования T- и B-лимфоцитов и иммуноцитов (например, плазмоцитов).
Миелоидная и лимфоидная ткани являются разновидностями соединительной ткани, т.е. относятся к тканям внутренней среды. В них представлены две основные клеточные линии — клетки ретикулярной ткани и гемопоэтические клетки.
Ретикулярные, а также жировые, тучные и остеогенные клетки вместе с межклеточным веществом формируют микроокружение для гемопоэтических элементов. Структуры микроокружения и гемопоэтические клетки функционируют в неразрывной связи друг с другом. Микроокружение оказывает воздействие на дифференцировку клеток крови (при контакте с их рецепторами или путем выделения специфических факторов).
Таким образом, для миелоидной и всех разновидностей лимфоидной ткани характерно наличие стромальных и гемопоэтических элементов, образующих единое функциональное целое.
СКК относятся к самоподдерживающейся популяции клеток. Они редко делятся. Выявление СКК стало возможным при применении метода образования клеточных колоний – потомков одной стволовой клетки.
Пролиферативную активность СКК регулируют колониестимулирующие факторы (КСФ), различные виды интерлейкинов (ИЛ-3 и др.). Каждая СКК в эксперименте или лабораторном исследовании образует одну колонию и называется колониеобразующей единицей (сокращенно КОЕ, CFU).
Исследование клеточного состава колоний позволило выявить две линии их дифференцировки. Одна линия дает начало мультипотентной клетке — родоначальнице гранулоцитарного, эритроцитарного, моноцитарного и мегакариоцитарного рядов гемопоэза (сокращенно КОЕ-ГЭММ). Вторая линия дает начало мультипотентной клетке — родоначальнице лимфопоэза (КОЕ-Л).
Из мультипотентных клеток дифференцируются олигопотентные (КОЕ-ГМ) и унипотентные родоначальные клетки. Методом колониеобразования определены родоначальные унипотентные клетки для моноцитов (КОЕ-М), нейтрофильных гранулоцитов (КОЕ-Гн), эозинофилов (КОЕ-Эо), базофилов (КОЕ-Б), эритроцитов (БОЕ-Э и КОЕ-Э), мегакариоцитов (КОЕ-МГЦ), из которых образуются клетки-предшественники. В лимфопоэтическом ряду выделяют унипотентные клетки — предшественницы для B-лимфоцитов и для T-лимфоцитов. Полипотентные (плюрипотентные и мультипотентные), олигопотентные и унипотентные клетки морфологически не различаются.
Все приведенные выше стадии развития клеток составляют четыре основных класса, или компартмента, гемопоэза:
- I класс — СКК - стволовые клетки крови (плюрипотентные, полипотентные);
- II класс — КОЕ-ГЭММ и КОЕ-Л - коммитированные мультипотентные клетки (миелопоэза или лимфопоэза);
- III класс — КОЕ-М , КОЕ-Б и т.д. - коммитированные олигопотентные и унипотентные клетки;
- IV класс — клетки-предшественники (бласты , напр.: эритробласт, мегакариобласт и т.д.).
Сразу отметим, что оставшиеся два класса гемопоэза составляют созревающие клетки (V класс) и зрелые клетки крови (VI класс).
Эритропоэз у млекопитающих и человека протекает в костном мозге в особых морфофункциональных ассоциациях, получивших название эритробластических островков. Эритробластический островок состоит из макрофага, окруженного одним или несколькими кольцами эритроидных клеток, развивающихся из унипотентной КОЕ-Э, вступившей в контакт с макрофагом. КОЕ-Э и образующиеся из нее клетки (от проэритробласта до ретикулоцита) удерживаются в контакте с макрофагом его рецепторами.
У взрослого организма потребность в эритроцитах обычно обеспечивается за счет усиленного размножения эритробластов. Но всякий раз, когда потребность организма в эритроцитах возрастает (например, при потере крови), эритробласты начинают развиваться из предшественников, а последние — из стволовых клеток.
В норме из костного мозга в кровь поступают только эритроциты и ретикулоциты.
Моноцитопоэз – процесс образования моноцитов, протекает в красном костном мозге и описывается последовательностью:
СКК -> КОЕ –ГЭММ -> КОЕ-ГМ -> КОЕ-М -> монобласт -> промоноцит -> моноцит
Промоноцит – сравнительно крупная клетка (диаметром 12-18 мкм) с большим светлым ядром, в котором располагается 1-2 ядрышка. Базофильная цитоплазма содкржит умеренно развитую грЭПС, полисомы, митохондрии, центриоли и крупный комплекс Гольджи, от которого отделяются незрелые азурофильные гранулы. Промоноциты делятся и постепенно дифференцируются в моноциты.
Моноциты покидают костный мозг вскоре после формирования, не образуя резервного костномозгового пула. Выделяясь в синусы костного мозга, они попадают в кровь, в которой циркулируют от 8ч до 3-4 суток, а затем через стенку сосудов мигрируют в ткани. Лишь около 5% моноцитов циркулируют в крови, остальные находятся во внесосудистом пуле. В тканях они превращаются в различные виды макрофагов и в дендритные антиген – представляющие клетки.
2.3.Регуляция гемопоэза
Кроветворение регулируется:
- факторами роста, обеспечивающими пролиферацию и дифференцировку СКК и последующих стадий их развития,
- факторами транскрипции, влияющими на экспрессию генов, определяющих направление дифференцировки гемопоэтических клеток,
- витаминами, гормонами.
Факторы роста включают колониестимулирующие факторы (КСФ), интерлейкины и ингибирующие факторы. Они являются гликопротеинами, действующими и как циркулирующие гормоны, и как местные медиаторы, регулирующие гемопоэз и дифференцировку специфических типов клеток. Почти все факторы роста действуют на СКК, КОЕ, коммитированные и зрелые клетки. Однако отмечаются индивидуальные особенности действия этих факторов на клетки-мишени.
КСФ действуют на специфические клетки или группы клеток на различных стадиях дифференцировки. Например, фактор роста стволовых клеток влияет на пролиферацию и миграцию СКК в эмбриогенезе. В постнатальном периоде на гемопоэз оказывают влияние несколько КСФ, среди которых наиболее изучены факторы, стимулирующие развитие гранулоцитов и макрофагов (ГМ-КСФ, Г-КСФ, М-КСФ), а также интерлейкины.
Большинство указанных факторов выделено и применяется для лечения различных болезней. Для получения их используются биотехнологические методы.
Дифференцировка полипотентных клеток в унипотентные определяется действием ряда специфических факторов, поэтинов — эритропоэтинов (для эритробластов), гранулопоэтинов (для миелобластов), лимфопоэтинов (для лимфобластов), тромбопоэтинов (для мегакариобластов).
Большая часть эритропоэтина образуется в почках. Его образование регулируется содержанием в крови кислорода, которое зависит от количества циркулирующих в крови эритроцитов. Снижение числа эритроцитов и соответственно парциального давления кислорода, является сигналом для увеличения продукции эритропоэтина. Эритропоэтин действует на чувствительные к нему КОЕ-Э, стимулируя их пролиферацию и дифференцировку, что в конечном итоге приводит к повышению содержания в крови эритроцитов.
Тромбопоэтин синтезируется в печени, стимулирует пролиферацию КОЕ-МГЦ, их дифференцировку и образование тромбоцитов.
Ингибирующие факторы дают противоположный эффект, т.е. тормозят гемопоэз; их недостаток может быть одной из причин лейкемии, характеризующейся значительным увеличением числа лейкоцитов в крови. Выделен ингибирующий лейкемию фактор (ЛИФ), который тормозит пролиферацию и дифференцировку моноцитов-макрофагов.
Витамины необходимы для стимуляции пролиферации и дифференцировки гемопоэтических клеток. Витамин В12 поступает с пищей и соединяется с внутренним фактором (Касла), который синтезируется париетальными клетками желудка. Образуемый при этом комплекс, в присутствии ионов Са2+, соединяется с рецепторами эпителиоцитов подвздошной кишки и всасывается. При всасывании в эпителиоциты поступает лишь витамин В12, а внутренний фактор освобождается. Витамин В12 поступает с кровью в костный мозг, где влияет на гемопоэз, и в печень, где может депонироваться. Нарушение процесса всасывания при различных заболеваниях желудочно-кишечного тракта может служить причиной дефицита витамина В12 и нарушений в гемопоэзе.
3. Моноциты
Моноциты - наиболее крупные (12-20 мкм) клетки крови. Форма ядра разнообразная, от круглой до неправильной с многочисленными выступами и углублениями поверхности. Хроматиновая сеть в ядре имеет рыхлое строение. Моноциты обладают резко выраженной способностью к окрашиванию, амебойдному движению и фагоцитозу, особенно остаток клеток, чужеродных мелких тел и т. д.
Моноциты составляют от 2 до 10 % всех лейкоцитов, способны к амебовидному движению, проявляют выраженную фагоцитарную и бактерицидную активность. Они фагоцитируют до 100 микробов, в то время как нейтрофилы - лишь 20-30. Моноциты появляются в очаге воспаления после нейтрофилов и проявляют максимум активности в кислой среде, в которой нейтрофилы теряют свою активность. В очаге воспаления моноциты фагоцитируют микробы, а также погибшие лейкоциты, поврежденные клетки воспаленной ткани, очищая очаги воспаления и подготавливая его для регенерации. За эту функцию моноциты называют дворниками организма.
Моноциты - самая большая клетка крови диаметром около 15 мкм. Поверхность ворсинчатая или гофрированная. У эмбрионов моноцит является первой клеткой, образующейся в желточном мешке. Выход моноцитов в кровь осуществляется около 72 часов. Общее количество циркулирующих моноцитов составляет 2 - 10% от всей популяции лейкоцитов. При этом только около 1% моноцитов крови пролиферируют и обладают выраженной фагоцитарной активностью.
Некоторые моноциты покидают кровь случайными, невыясненными путями, а большинство - путем адгезии на стенках синусов в печени, селезенке и железах внутренней секреции. Данная популяция моноцитов взаимодействует с подлежащими эпителиальными клетками и является идеальным "стражем" этих органов от патогенов. Моноциты проходят стенку капилляров путем диапедеза и устремляются в очаги поражения.
При переходе моноцитов из крови в ткани, большая их часть подвергается апоптозу, вследствие отсутствия в тканевом микроокружении необходимых факторов защиты от апоптоза.
Моноциты трансформируются в макрофаги, увеличиваясь в размерах в 5-10 раз, диаметром 25-50 мкм. Клетки с большим количеством ламеллоподий и микроворсинок. При этом резко возрастает фагоцитарная активность, снижается содержание гидролитических ферментов в лизосомах, уменьшается количество митохондрий. В тканях резидентные макрофаги могут находиться до нескольких месяцев и вынуждены выбирать между двумя возможностями: либо стать "фиксированными", либо вновь трансформироваться в блуждающую клетку.
Моноциты, составляющие около 5% лейкоцитов крови, находятся в циркуляции около 1 сут., а затем поступают в ткани, формируя популяцию тканевых макрофагов, количество которых в 25 больше, чем моноцитов. К ним относятся купферовские клетки печени, микроглия центральной нервной системы, остеокласты костной ткани, макрофаги легочных альвеол, кожи и других тканей. Много макрофагов во всех органах иммунной системы. Тканевые макрофаги - клетки с округлым или почковидным ядром имеют диаметр 40-50 мкм. Цитоплазма содержит лизосомы с набором гидролитических ферментов, обеспечивающих переваривание любых органических веществ и выделение бактерицидного аниона кислорода. Макрофаги функционируют как фагоциты. Они продуцируют растворимые вещества, регулирующие другие клетки иммунной системы, из которых наиболее изучен ИЛ-1, активирующий лимфоциты. На мембране макрофага экспрессированы структуры, обеспечивающие способность отличать чужеродные субстраты от собственных. Маркер макрофага - белок СОН служит рецептором липополисахаридов бактерий. Макрофаг обладает пектиноподобными молекулами, соединяющимися с маннозными и фруктозными компонентами поверхности большинства микроорганизмов, что
9-09-2015, 00:12