Концентрацію газу в крові характеризує його напруження – тиск газу, розчиненого в рідині. Напруження також позначається літерою Р.
Газовий склад вдихуваного (атмосферного), видихуваного та альвеолярного повітря різний:
| Повітря | О2 | СО2 | Азот та ін. гази. |
| Атмосферний | 20,93% | 0,03% | 79,04% |
| Видихуваний | 16,0% | 4,5% | 79,5% |
| Альвеолярний | 14,0% | 5,5% | 80,5% |
Причиною різного газового складу атмосферного та видихуваного повітря є газообмін в легенях. Причиною різного газового складу видихуваного та альвеолярного повітря являється те, що видихуване повітря містить окрім альвеолярного ще й повітря із дихальних шляхів (мертвий простір, який не відрізняється за складом від атмосферного).
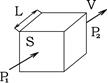 Дифузія газів в легенях підкоряється закону Фіка: об’єм дифузії газу (V) прямо пропорційний площі дифузії (S), коефіцієнту дифузії (К), градієнту тиску газу по обидві сторони альвеоло-капілярної мембрани (Р1
– Р2
) і обернено пропорційний товщині цієї мембрани (L):
Дифузія газів в легенях підкоряється закону Фіка: об’єм дифузії газу (V) прямо пропорційний площі дифузії (S), коефіцієнту дифузії (К), градієнту тиску газу по обидві сторони альвеоло-капілярної мембрани (Р1
– Р2
) і обернено пропорційний товщині цієї мембрани (L):
![]()
Площа дифузії в легенях ( S ) – це площа альвеол, які вентилюються та кровопостачаються. При різних функціональних станах у дорослої людини цей показник змінюється від 80 до 120 м2 . Збільшення площі дифузії може зумовити збільшення глибини дихання і об’ємної швидкості кровотоку через судини легень, що має місце, наприклад, при фізичному навантаженні.
Товщина дифузного шару ( L ) у здорової людини складає 0,4 – 1,5 мкм. Дифузний шар (альвеоло-капілярна мембрана) включає в себе: шар сурфактанту, шар пневмоцитів, базальну мембрану альвеоли, шар ніжної сполучної тканини, базальну мембрану капіляра, шар ендотеліоцитів. В фізіологічних умовах товщина дифузного шару зменшується при збільшенні глибини дихання.
Коефіцієнт дифузії (К) є різним для різних газів і визначається їх здатністю розчинятися в структурах, які формують дифузний шар. За хімічним складом в цих структурах переважають фосфоліпіди (сурфактант, мембрани пневмоцитів та ендотеліоцитів, базальні мембрани) і вода. СО2 в фосфоліпідах та воді розчиняється набагато краще, ніж О2 , тому коефіцієнт дифузії для вуглекислого газу приблизно в 20 разів більший, ніж для кисню.
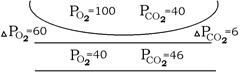 Градієнт тисків (Р1
– Р2
)
в стані спокою складає близько 60 мм.рт.ст. для О2
(100 – 40) та близько 6 мм.рт.ст. для СО2
(46 – 40). Оскільки Ро2
та Рсо2
в альвеолах підтримуються на постійному рівні, градієнт тисків цих газів змінюються за рахунок зміни їх напруження в венозній крові.
Градієнт тисків (Р1
– Р2
)
в стані спокою складає близько 60 мм.рт.ст. для О2
(100 – 40) та близько 6 мм.рт.ст. для СО2
(46 – 40). Оскільки Ро2
та Рсо2
в альвеолах підтримуються на постійному рівні, градієнт тисків цих газів змінюються за рахунок зміни їх напруження в венозній крові.
Таким чином, в стані спокою градієнт тисків, за яким йде дифузія О2 , в 10 разів більше градієнта, за яким іде дифузія СО2 . Та об’єм дифузії цих газів відрізняється на незначну кількість. Протягом однієї хвилини в стані спокою дифундує 250 – 300 мл. О2 і лише дещо менше СО2 (200 – 250 мл). Великий об’єм дифузії СО2 при порівняно невисокому градієнту тисків пов’язаний з високим коефіцієнтом дифузії цього газу.
Об’єм дифузії О2 і СО2 при фізичному навантаженні збільшується до десяти разів в порівнянні з станом спокою. Це пов’язано з збільшенням площі дифузії та зменшенням товщини дифузного шару, з збільшенням градієнту тисків газів по обидві сторони альвеоло-капілярної мембрани (за рахунок зміни газового складу венозної крові).
В клініці та експерименті за життя неможливо визначити площу дифузії і товщину дифузного шару. Тому для характеристики процесів дифузії в легенях ввели поняття дифузної здатності легень (ДЗЛ) – об’єм газу, який дифундує через альвеоло-капілярну мембрану за 1 хвилину при градієнті тиску цього газу по обидві сторони альвеоло-капілярної мембрани, що рівний 1 мм.рт.ст. Таким чином, визначають ДЗЛ шляхом поділу об’єму дифузії газу за одну хвилину на градієнт тисків цього газу в альвеолах і в крові:
![]()
ДЗЛ прямопропорційна площі дифузії, коефіцієнту дифузії даного газу, оберненопропорційна товщині дифузного шару і не залежить від градієнту тисків газу (дане визначення випливає, якщо замість V в формулі для визначення ДЗЛ підставити формулу з закону Фіка для визначення об’єму дифузії газу). ДЗЛ для О2
в стані спокою у дорослої людини складає близько 25 – 30 мл/хв ![]() мм.рт.ст.
мм.рт.ст.
В стані спокою час, протягом якого кров знаходиться в капілярах легень, складає 0,75 – 1,0с. Час, протягом якого проходить вирівнювання вмісту О2 в альвеолярному повітрі і в крові – близько 0,25с. Тобто, вміст газів в крові, відтікаючої від альвеоли, дорівнює такому в альвеолі. Така ж ситуація зберігається і при фізичному навантаженні – час проходження крові через капіляри легень скорочується до 0,25с, але все ж залишається достатнім для вирівнювання вмісту газів в крові та в альвеолярному повітрі. Тільки при дуже значному підвищенні лінійної швидкості руху крові в капілярах, її газовий склад не встигає досягнути параметрів альвеолярного повітря.
Та навіть в стані спокою має місце альвеоло-артеріальний градієнт тиску для О2 та СО2 – напруження кисню в артеріальній крові на 5 – 10 мм.рт.ст. нижче, а напруження вуглекислого газу на 1 – 2 мм.рт.ст. вище, ніж в альвеолярному повітрі (Ро2 = 100 мм.рт.ст., Рсо2 = 40 мм.рт.ст. в альвеолах; Ро2 = 90 – 95 мм.рт.ст., Рсо2 = 41 - 42 мм.рт.ст. в артеріальній крові). Причиною наявності альвеоло-артеріального градієнту тисків газів являється:
1. Наявність артеріо-венозних шунтів, по яким в артеріальну кров “скидається” венозна. В якості таких шунтів виступають:
- судини, що забезпечують живлення бронхіального дерева і легень;
- вени Тебехія, по яким венозна кров від серця надходить в ліві його відділи;
- капіляри, за якими рухається кров через невентильовані альвеоли.
2. Нерівномірність вентиляції і перфузії (кровотоку) в легенях. Для легенів в цілому відношення вентиляції до перфузії (АВЛ до ХОК) складає близько 0,8:
![]() ;
;
При такому рівні ВПВ (вентиляційно-перфузійних відношень) Ро2 в альвеолах рівняється 100 мм.рт.ст., а Рсо2 – 40 мм.рт.ст. Та такий рівень ВПВ має місце тільки в альвеолах середніх сегментів легень, в верхівках і в базальних сегментах легень ВПВ інше.
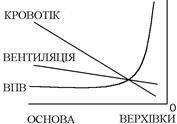 Причина полягає в тому, що в легенях має місце:
Причина полягає в тому, що в легенях має місце:
а) нерівномірність вентиляції: краще всього вентилюються базальні сегменти легень, менше – середні сегменти, найбільш погано – верхівки легень. Причина нерівномірності вентиляції заключається в тому, що верхівки легень розтягнені силою тяжіння вже в стані видиху, тому їх здатність до розтягнення нижча і на той же приріст транспульмонального тиску вони менше збільшують свій об’єм Ù гірше вентилюються.
б) нерівномірність перфузії: артеріальні судини легень мають дуже високу здатність до розтягнення (як і венозні), тому кровотік в них в значній мірі залежить від впливу фактора гравітації – від серця основна маса крові під впливом сили тяжіння направляється до базальних сегментів легень, менша – до середніх, сама мала – до верхівок.
Та ступінь нерівномірності вентиляції і кровотоку різна. Нерівномірність кровотоку виражена набагато більше, ніж нерівномірність вентиляції. Тому ВПВ змінюється в нвпрямку від верхівок до базальних сегментів легень:
- в верхівках знижена вентиляція і кровотік, та більша ступінь зниження кровотоку призводить до підвищення ВПВ Ù переважання вентиляції Ù зміна газового складу альвеолярного повітря (збільшення Ро2 і зниження Рсо2 ) Ù аналогічна зміна газового складу відтікаючої від цих альвеол крові, тобто, кров, відтікаюча від верхівок, має Ро2 більше 100 мм.рт.ст., а Рсо2 менше 40 мм.рт.ст.;
- в середніх сегментах легень ВПВ = 0,8 Ù Ро2 в альвеолах (і крові) складає 100 мм.рт.ст., Рсо2 – 40 мм.рт.ст.;
- в базальних сегментах легень переважає кровотік, ВПВ = 0,7 – 0,6, тому Ро2 в альвеолах (і крові) менше 100 мм.рт.ст., а Рсо2 – більше 40 мм.рт.ст.
Всі роздуми, що були наведенні вище, торкаються вертикальної пози людини. Оскільки при цьому основна маса крові направляється до базальних сегментів легень, де ВПВ = 0,8, в артеріальній крові, що відтікає від легень до лівого серця Ро2 = 100 мм.рт.ст., а Рсо2 = 40 мм.рт.ст.
Внесок названих факторів (артеріо-венозного шунтування та нерівномірності ВПВ) в створення альвеоло-артеріальних градієнтів Ро2 та Рсо2 приблизно рівний.
6. Транспорт кисню кров’ю. Киснева ємкість крові.
Кисень транспортується кров’ю у двох формах:
1. Розчинений у плазмі крові. При РСО2 =100 мм.рт.ст. в 1л крові розчиняється 3 мл кисню.
2. В хімічно зв’язаному з гемоглобіном стані – у вигляді оксигемоглобіну. Це основна форма транспорту кисню – 1г гемоглобіну за оптимальних умов може зв’язати 1,34 мл кисню. Виходячи з цього розраховують кисневу ємкість крові – максимальну кількість О2 , котру може зв’язати 1л крові. КЄК при концентрації гемоглобіну 150 г/л складає 200мл/л, або 20% об’ємних.
7. Крива дисоціації оксигемоглобіну, фактори, що впливають
на її хід.
Здатність гемоглобіну реагувати з киснем характеризує крива дисоціації оксигемоглобіну (КДО). При її побудові на осі абсцис відкладують РО2 (мм.рт.ст), по осі ординат – відсотковий вміст оксигемоглобіну в крові. Будують КДО мінімум за двома точками:
- при РО2 = 100 мм.рт.ст. гемоглобін насичений киснем на 98%;
- при РО2 = 60 мм.рт.ст. насичення О2 складає 90%;
- при РО2 = 26 мм.рт.ст. насичення гемоглобіну киснем – 50%.
- при РО2 = 0, насичення гемоглобіну = 0%.
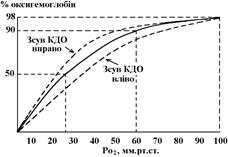 Характерно, що при високому РО2
гемоглобін легко взаємодіє з киснем – утворення HbO2
(верхня, полога – "горизонтальна" частина кривої). Зниження РО2
зі 100 до 60 мм.рт.ст мало впливає на утворення НbO2
– його концентрації зменшується лише на 8%. Це означає, що зниження тиску кисню в альвеолах до 60 мм.рт.ст мало вплине на транспорт кисню кров’ю, хоча напруження кисню в плазмі буде знижуватися пропорційно зниженню тиску О2
в альвеолах. Завдяки такій особливості ходу КДО, ми можемо, наприклад, підійматися в гори – не зважаючи на істотне зниження атмосферного тиску, постачання тканин киснем зберігається на потрібному рівні.
Характерно, що при високому РО2
гемоглобін легко взаємодіє з киснем – утворення HbO2
(верхня, полога – "горизонтальна" частина кривої). Зниження РО2
зі 100 до 60 мм.рт.ст мало впливає на утворення НbO2
– його концентрації зменшується лише на 8%. Це означає, що зниження тиску кисню в альвеолах до 60 мм.рт.ст мало вплине на транспорт кисню кров’ю, хоча напруження кисню в плазмі буде знижуватися пропорційно зниженню тиску О2
в альвеолах. Завдяки такій особливості ходу КДО, ми можемо, наприклад, підійматися в гори – не зважаючи на істотне зниження атмосферного тиску, постачання тканин киснем зберігається на потрібному рівні.
Коли парціальний тиск О2 в атмосфері високий, реакція
Hb + O2 = HbO2
зсунута в бік утворення оксигемоглобіну. В умовах цілісного організму такі умовистворюються при проходженні крові капілярами легень.
Зниження Ро2 нижче 60 мм.рт.ст. супроводжується значним зниженням HbO2 в крові – він активно дисоціює з утворенням гемоглобіну та вільного кисню. В умовах цілісного організму це відбувається в тканинах (рівень Ро2 складає 50-20 мм.рт.ст.). І що активніше функціонує тканина, тим нижчий в ній рівень О2 – посилена дисоціація HbO2 з вивільненням молекулярного кисню, котрий утилізується тканинами. Тобто, за цих умов реакція взаємодії кисню та гемоглобіну зсунута в бік дисоціації оксигемоглобіну.
Отже, S-подібна форма кривої дисоціації оксигемоглобіну відображає компроміс між необхідністю активно зв’язувати кисень у атмосфері з високим парціальним тиском його (капіляри легень) та легко віддавати кисень в атмосфері з низьким парціальним тиском (капіляри тканин).
Спорідненість кисню та гемоглобіну і хід КДО залежать від кількох чинників. Деякі з них знижують цю спорідненість; прицьому гемоглобін легше віддає кисень, а КДО зсувається праворуч. Видно, що при зсуві КДО праворуч при тому ж рівні парціального тиску кисню розпадається більше оксигемоглобіну. До чинників, що зсувають КДО праворуч належать:
- накопичення вуглекислоти (взаємодіє з глобіновою частиною Нb – зниження його спорідненості до кисню);
- накопичення йонів водню (протонів – зниження рН) – протони також взаємодіють з глобіном – зниження спорідненості кисню і Hb;
- підвищення температури.
Усі ці цинники впливають на оксигемоглобін в тканинах, що активпо працюють – саме там накопичуються вуглекислота, протони, підвищується температура (через посилений метаболізм). Це призводить до зменшення спорідненості Hb і О2 – посилення дисоціації оксигемоглобіну – посилене утворення молекулярного кисню, котрий необхідний тканинам, що активно функціонують.
Ще одним чинником, котрий зсуває КДО праворуч є підвищення концентрації 2,3-дифосфогліцерату (2,3-ДФГ) в еритроцитах. Ця сполука є продуктом вуглеводного обміну (гліколіз). А оскільки основним джерелом енергії для еритроцитів є анаеробний гліколіз, то 2,3-ДФГ виникає при гіпоксії. 2,3-ДФГ взаємодіє з глобіном – спричиняє аналогічний з попередніми сполуками ефект – покращує постачання тканин киснем.
Зсув КДО ліворуч відповідає підвищенню спорідненості гемоглобіну до кисню – при тому ж рівні парціального тиску О2 кількість оксигемоглобіну буде більшою. Зсув КДО ліворуч викликають:
- зниження концентрації вуглекислоти;
- підвищення рН;
- зниження температури;
- зниження вмісту 2,3-ДФГ в еритроцитах.
Отже, чинниками, котрі впливають на зв’язування і передачу кисню кров’ю є:
- вміст гемоглобіну в крові (прямопропорційний до зв’язувальної здатності крові);
- парціальний тиск кисню (при високому тиску переважно зв’язує кисень, при високому - віддає);
- вміст вуглекислоти;
- рН;
- температура;
- концентрація 2,3-ДФГ;
Коефіцієнт утилізації кисню (КУО2 ) характеризує віддачу кисню тканинам. Розраховують його за формулою:
![]() Виражають КУО2
в процентах. У стані спокою він складає 30-35%, тобто тканини споживають 30-35% кисню, що міститься в артеріальній крові. При фізичному навантаженні КУО2
збільшується; при дуже інтенсивній роботі він складає 70-75%. Цьому сприяють:
Виражають КУО2
в процентах. У стані спокою він складає 30-35%, тобто тканини споживають 30-35% кисню, що міститься в артеріальній крові. При фізичному навантаженні КУО2
збільшується; при дуже інтенсивній роботі він складає 70-75%. Цьому сприяють:
- зниження Ро2 в тканинах, що активно функціонують;
- накопичення в цих тканинах вуглекислоти;
- накопичення в них протонів;
- підвищення в них температури.
Завдяки підвищенню утилізації кисню при фізичному навантаженні зростає артеріо-венозна різниця кисню. У спокої вміст О2 в артеріальній кровіблизько 200 мл/л, у венозній – 130-140 мл/л. Тобто артеріо-венозна рязниця складає 60-70мл/л. При фізичному навантаженні вміст О2 в артеріальній крові зростає незначно (вихід крові з депо і згущеня її в одиниці об’єму дещо підвищує концентрацію гемоглобіну). В той же час істотно знижується вміст О2 у венозній крові за рахунок посиленої його віддачі; вміст О2 прицьому у венозній крові може знижуватися до 60-70 мл/л. За таких умов артеріо-венозна різниця зростає до 130-140 мл/л.
8. Транспорт вуглекислого газу кров'ю. Роль еритроцитів в транспорті вуглекислого газу.
Вуглекислий газ транспортується наступними шляхами:
1. Розчинений у плазмі крові – близько 25 мл/л.
2. Зв’язаний з гемоглобіном (карбгемоглобін) – 45 мл/л.
3. У вигляді солей вугільної кислоти – букарбонати каліі та натрію плазми крові – 510 мл/л.
Таким чином, у стані спокою кров транспортує 580 мл вуглекислого газу в 1 л. Отже, основною формою транспорту СО2 є бікорбонати плазми, що утворюються завдяки активному протіканню карбоангідразної реакції.
В еритроцитах міститься фермент карбоангідраза (КГ), котрий каталізує взаємодію вуглекислого газу із водою з утворенням вугільної кислоти, що дисоціює з утворенням бікарбонатного йона та протона. Бікарбонат всередині еритроцита взаємодіє з йонами калію, що виділяються з калієвої солі гемоглобіну при відновленні останнього. Так всередині 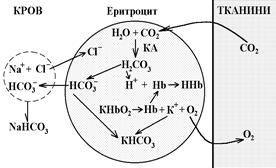 еритроцита утворюється бікарбонат калію. Але бікарбонатні йони утворюються в значній концентрації і тому за градієнтом концентрації (в обмін на йони хлору) надходять у плазму крові. Так у плазмі утворюється бікарбонат натрію. Протон, що утворився при дисоціації вугільної кислоти, реагує з гемоглобіном з утворенням слабкої кислоти ННb.
еритроцита утворюється бікарбонат калію. Але бікарбонатні йони утворюються в значній концентрації і тому за градієнтом концентрації (в обмін на йони хлору) надходять у плазму крові. Так у плазмі утворюється бікарбонат натрію. Протон, що утворився при дисоціації вугільної кислоти, реагує з гемоглобіном з утворенням слабкої кислоти ННb.
В капілярах легень ці процеси йдуть в зворотньому напрямку. З йонів водню та бікарбонатних йонів утворюється вугільна кислота, котра швидко розпадається на вуглекислий газ та воду. Вуглекислий газ видаляється назовні.
Отже, роль еритроцитів у транспорті вуглекислоти така:
- утворення солей вугільної кислоти;
- утворення карбгемоглобіну.
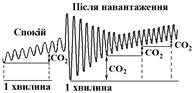 Дифузія газів в тканинах підкоряється загальним законам (об’єм дифузії прямопропорційний площі дифузії, градієнту напруження газів в крові та тканинах). Площа дифузії збільшується, а товщина дифузного шару зменшується під час збільшення кількості функціонуючих капілярів, що має місце при підвищенні рівня функціональної активності тканин. В цих же умовах зростає градієнт напруження газів за рахунок зниження в активно працюючих органах Ро2
та підвищення Рсо2
(газовий склад артеріальної крові, як і альвеолярного повітря залишається незмінним!!!). Всі ці зміни в активно працюючих тканинах сприяють збільшенню об’єму дифузії О2
та СО2
в них. Споживання О2
(СО2
) за спірограмою визначають по зміні (зсуву) кривої вверх за одиницю часу (1 хвилину).
Дифузія газів в тканинах підкоряється загальним законам (об’єм дифузії прямопропорційний площі дифузії, градієнту напруження газів в крові та тканинах). Площа дифузії збільшується, а товщина дифузного шару зменшується під час збільшення кількості функціонуючих капілярів, що має місце при підвищенні рівня функціональної активності тканин. В цих же умовах зростає градієнт напруження газів за рахунок зниження в активно працюючих органах Ро2
та підвищення Рсо2
(газовий склад артеріальної крові, як і альвеолярного повітря залишається незмінним!!!). Всі ці зміни в активно працюючих тканинах сприяють збільшенню об’єму дифузії О2
та СО2
в них. Споживання О2
(СО2
) за спірограмою визначають по зміні (зсуву) кривої вверх за одиницю часу (1 хвилину).
9. Фізіологічна роль дихальних шляхів, регуляція їх просвіту.
Дихальні шляхи виконують дуже важливу функцію – кондиціювання повітря. Завдяки цьому в легені надходить повітря тільки певних параметрів. В дихальних шляхах повітря:
- зігрівається;
- зволожується, тому повітря в легенях насичене водяними парами на 100%, незалежно від вологості атмосферного повітря;
- очищується, завдяки наявності війчастого епітелію та бокалоподібних клітин, які секретують слиз (рух війок забезпечує проходження слизу і осівших на поверхні дихальних шляхів чужорідних частинок, мікроорганізмів в напрямку гортані та глотки, де вони проковтуються або відхаркуються), частина осівших на поверхні дихальних шляхів мікроорганізмів і частинок знешкоджуються макрофагами.
Окрім того, повітроносні шляхи мають велику кількість рецепторів (є рефлексогенною зоною), що забезпечує їх участь в здійсненні захисних дихальних рефлексів.
В регуляції величини просвіту дихальних шляхів (і їх опору руху повітря) приймають участь, головним чином, нервові механізми. При цьому парасимпатичні рефлекторні впливи супроводжуються скороченням гладких м’язів Ù звуження дихальних шляхів. В гладких м’язах дихальних шляхів є a- та b-адренорецептори; стимуляція b-адренорецепторів супроводжується розслабленням гладких м’язів повітроносних шляхів і розширенням бронхіол, а стимуляція a-адренорецепторів – напруженям гладких м’язів та звуженням бронхів. В гладких м’язах дихальних шляхів переважають b-адренорецептори, тому в умовах підвищення активності симпато-адреналової системи просвіт дихальних шляхів розширюється Ù створюються оптимальні умови для вентиляції легень.
10. Дихальний центр, його будова, регуляція ритмічності дихання.
Регуляція зовнішнього дихання здійснюється шляхом ритмічного чергування вдиху та видиху. Вдих та видих проходять завдяки скорочення та розслаблення скелетних м’язів. Скорочення та розслаблення їх регулюється тільки за допомогою нервових механізмів (соматичних рефлексів). Зміна
8-09-2015, 23:15