Дихальний центр (ДЦ) знаходиться в задньому мозку (довгастий мозок та міст), хоча в регуляції зовнішнього жихання приймають участь всі рівні ЦНС, аж до кори головного мозку. Локалізація ДЦ визначається в дослідах з перерізкою стовбуру мозку.
Таким чином, перерізка стовбуру вище заднього мозку не супроводжується змінами дихання, а перерізка нижче довгастого мозку призводить до його зупинки Ù ДЦ знаходиться саме в задньому мозку. Та він має складну структуру, тому що перерізки на різних рівнях заднього мозку призводить до різних порушень дихальної ритміки (апнейзіс – глибоке дихання з затримкою на вдосі; гаспінг – глибоке дихання з затримкою на видосі). В верхній третині мосту знаходиться пневмотаксичний центр (ПТЦ), який припиняє вдих та забезпечує поступовий перехід вдиху до видиху (перерізка стовбура на цьому рівні призводить до розвитку апнейстичного дихання).
Дихальний центр довгастого мозку має два ядра – дорсальне та вентральне (розміщені симетрично по відношенню до середньої лінії).
Особливістю дорсального ядра є наявність в ньому тільки інспіраторних нейронів, які збуджуються безпосередньо перед вдихом та під час вдиху (інспіраторні нейрони або нейрони вдиху). Нейрони дорсального дихального ядра забезпечують регуляцію дихання в стані спокою (при цьому акт вдиху є активним, а видиху – пасивним). Збудження інспіраторних нейронів дорсального ядра забезпечує скорочення м’язів спокійного вдиху Ù вдих Ù гальмування інспіраторних нейронів дорсального ядра Ù розслаблення м’язів Ù пасивний видих.
До складу вентрального ядра входять як інспіраторні так і експіраторні нейрони (нейрони видиху). Це ядро вмикається в регуляцію форсованого (глибокого): дихання інспіраторні нейрони вентрального ядра забезпечують скорочення м’язів форсованого вдоху; експіраторні – м’язів видиху.
Еферентні зв’язки нейронів дорсального ядра (інспіраторних) здійснюються таким чином: від цих нейронів по ретикулоспінальних шляхах інформація передається до мотонейронів спинного мозку:
- які локалізуються в ІІІ – ІV сегментах шийного відділу спинного мозку; аксони цих нейронів формують діафрагмальний нерв Ù передача інформації до діафрагми;
- які локалізуються в усіх сегментах грудного відділу Ù до зовнішніх міжреберних м’язів.
Інспіраторні нейрони вентрального дихального ядра передають інформацію до мотонейронів спинного мозку, які інервують додаткові м’язи вдиху, експіраторні – до мотонейронів, які інервують м’язи видиху.
Механізм ритмічного чергування вдиху та видиху в стані спокою пов’язаний з ритмічним збдженням та гальмуванням інспіраторних нейронів дорсального ядра дихального центру.
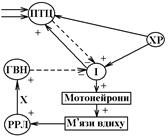 Збудження цих нейронів головним чином зумовлюється надходженням до них інформації від хеморецепторів (ХР) (центральних і периферичних), які є тонічно активними: генерують ПД навіть при повністю нормальному газовому складу артеріальної крові та ліквора. Збудження від цих інспіраторних нейронів передається до мотонейронів спинного мозку Ù збудження та скорочення дихальних м’язів Ù вдих.
Збудження цих нейронів головним чином зумовлюється надходженням до них інформації від хеморецепторів (ХР) (центральних і периферичних), які є тонічно активними: генерують ПД навіть при повністю нормальному газовому складу артеріальної крові та ліквора. Збудження від цих інспіраторних нейронів передається до мотонейронів спинного мозку Ù збудження та скорочення дихальних м’язів Ù вдих.
Далі збудження інспіраторних нейронів дорсального ядра змінюється їх гальмуванням. Цьому сприяє надходження до них інформації від таких структур:
1. Від рецепторів розтягнення легень (РРЛ) по аферентних волокнах блукаючого нерва Ù спочатку до гальмівних вставних нейронів (ГВН) Ù до інспіраторних нейронів;
2. Від пневмотаксичного центру (ПТЦ) , збудження якого при вдосі пов’язане з надходженням до нього інформації від інспіраторних нейронів, а також від інших нейронів (ПТЦ – частина ретикулярної формації стовбура мозку і його аферентні зв’язки різноманітні). Коли потік гальмівної інформації від РРЛ та від ПТЦ до інспіраторних нейронів стає більш потужнішим, ніж потік збуджуючої інформації від хеморецепторів, інспіраторні нейрони гальмуються Ù гальмування мотонейронів спинного мозку Ù розслаблення дихальних м’язів Ù видих.
Видих знімає механізми, які активують РРЛ та ПТЦ Ù припинення надходження гальмівної інформації до інспіраторних нейронів Ù переважання аферентного входу від ХР Ù збудження інспіраторних нейронів Ù видих.
11. Механізм першого вдиху новонародженої дитини.
1. Після перерізки пуповини в крові дитини накопичується вуглекислота, знижується рН крові та знижується парціальний тиск кисню – стимуляція центральних та периферичних хеморецепторів – збудження дихального центру – збудження інспіраторних нейронів – скорочення м’язів вдиху.
2. Стимулювати перший вдих новонародженого можна дією на різні рецептори, оскільки аферентна інервація дихального центру дуже різноманітна:
- стимулюючи терморецептори – раніше дитину опускали то в холодну то в теплу воду;
- діючи на механорецептори (поплескування, пощипування шкіри);
- діючи на вестибулорецептори – дитину можна опускати головою вниз, нахиляти зі сторони в сторону.
3. Після народження дитини треба зняти “рефлекс пірнальника”, котрий блокує настання вдиху через наявність рідини в дихальних шляхах. Щоб зняти цей рефлекс цю рідину періодично аспірують.
12. Роль рецепторів розтягнення легень та блукаючих нервів в регуляції дихання.
Рецептори розтягнення легень збуджуються у відповідь на розтягнення легенів при вдосі, особливо при глибокому. Інформація від цих рецепторів передається в дихальний центр по аферентних волокнах блукаючого нерва і через гальмівні вставні нейрони гальмує інспіраторні нейрони Ù видих. Таким чином, за участю цих рефлексів, які починаються з подразнення цих рецепторів при вдосі, вдих припиняється і наступає видих. Ці рефлекси носять назву рефлексів Герінга-Брайера. Вони приймають участь в саморегуляції ритму дихання. Цим і визначається роль блукаючих нервів в забезпеченні ритму дихання.
Варто відзначити, що до рецепторів, які знаходяться в легенях та в дихальних шляхах, і які беруть участь в регуляції дихання відносяться:
- ірритантні рецептори легень , які реагують на дію їдких газів, пилу, тютюнового диму, холодного повітря і при збудженні зумовлюють звуження бронхів і гіпервентиляцію;
- юкстакапілярні рецептори , що розміщуються поблизу капілярів легень і реагують на зміну механічних властивостей легеневої тканини (набряк, переповнення капілярів кров’ю і т.д.) Ù тахіпное;
- рецептори дихальних шляхів , що реагують на механічні та хімічні подразники і спричиняють виникнення захисних дихальних рефлексів (кашель, чхання, звуження бронхів).
- пропріорецептори працюючих м’язів, інформація від яких стимулює зовнішнє дихання (рефлекторно) при фізичному навантаженні;
- пропріорецептори дихальних м’язів – забезпечують рефлекторну саморегуляцію дихання при зміні опору (на вдиху чи на видиху).
- артеріальні пресорецептори , які реагують на підвищення САТ Ù гіповентиляція. Протилежні реакції спостерігаються при зниженні САТ;
- вольові температурні рецептори – їх стимуляція призводить до різноманітних змін в акті дихання.
13. Роль центральних і периферичних хеморецепторів в регуляції дихання. Компоненти крові, що стимулюють зовнішнє дихання.
Дуже важливим є аферентий вплив на ДЦ хеморецепторів, які поділяються на центральні та периферичні:
Периферичні хеморецептори локалізуються в судинних рефлексогенних зонах (дуга аорти і каротидні синуси). В регуляції дихання більш важливими є каротидні хеморецептори. Адекватні подразники для них:
- збільшення Рсо2 артеріальної крові;
- зменшення рН артеріальної крові;
- зменшення Ро2 артеріальної крові.
Інформація, що надходить до дихального центру при підвищенні активності цих рецепторів, викликає гіпервентиляцію (підвищення глибини та частоти дихання) Ù нормалізація вказаних показників крові. Тобто, за їх участю здійснюється регуляція газового складу артеріальної крові за відхиленням (саморегуляція на основі негативного зворотнього зв’язку).
Центральні хеморецептори локалізуються на вентральній поверхні довгастого мозку (дно ІV шлуночка) і омиваються ліквором. Адекватними подразниками для цих рецепторів є:
- збільшення Рсо2 ліквора;
- зменшення рН ліквора.
Підвищення активності цих рецепторів, як і периферичних, призводить до розвитку гіпервентиляції Ù нормалізація вказаних показників ліквора.
Таким чином, периферичні хеморецептори, на відміну від центральних реагують на зниження Ро2 артеріальної крові. Тому гіпоксія стимулює зовнішнє дихання тільки за рахунок рефлексів, які розпочинаються з подразнення периферичних хеморецепторів. Та в цілому в регуляції зовнішнього дихання велике значення мають рефлекси, що пов’язані зі стимуляцією центральних хеморецепторів (співвідношення ролі центральних та периферичних хеморецепторів складає приблизно 4 : 1). Однак зміна Рсо2 і рН ліквора виникають пізніше, ніж артеріальної крові. Тому рефлекси з центральних хеморецепторів вмикаються в регуляцію зовнішнього дихання пізніше, ніж рефлекси з периферичних хеморецепторів.
Таким чином, не дивлячись на велику роль в регуляції зовнішнього дихання центральних хеморецепторів, периферичні хеморецептори:
- забезпечують більш швидкі рефлекторні відповіді;
- тільки вони реагують на гіпоксію Ù стимуляція зовнішнього дихання в умовах гіпоксії.
До компонентів крові, які стимулюють зовнішнє дихання, відносяться Ро2 , рН, Рсо2 . Найбільшого значення для регуляції зовнішнього дихання має вміст в крові СО2 (Рсо2 ), менше – вміст йонів Н+ , саме менше – вміст О2 (Ро2 ).
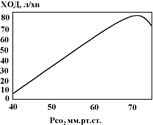 Залежність вентиляції легень від Рсо2
досліджують при диханні газовою сумішшю, в якій Ро2
залишається незмінним а Рсо2
поступово підвищується. При цьому відмічено, що підвищення Рсо2
в газовій суміші на 1 мм.рт.ст. супроводжується підвищенням ХОД на 2 – 3л/хв. При підвищенні Рсо2
артеріальної крові від 40 до 60 мм.рт.ст. ХОД зростає від 7 до 65 л/хв. Підвищення ХОД у відповідь на підвищення Рсо2
проходить лише до певної межі. Якщо Рсо2
в артеріальній крові стає вищим від 70 мм.рт.ст., ХОД знижується, що є результатом прямого пригнічуючого впливу СО2
на нейрони дихального центру.
Залежність вентиляції легень від Рсо2
досліджують при диханні газовою сумішшю, в якій Ро2
залишається незмінним а Рсо2
поступово підвищується. При цьому відмічено, що підвищення Рсо2
в газовій суміші на 1 мм.рт.ст. супроводжується підвищенням ХОД на 2 – 3л/хв. При підвищенні Рсо2
артеріальної крові від 40 до 60 мм.рт.ст. ХОД зростає від 7 до 65 л/хв. Підвищення ХОД у відповідь на підвищення Рсо2
проходить лише до певної межі. Якщо Рсо2
в артеріальній крові стає вищим від 70 мм.рт.ст., ХОД знижується, що є результатом прямого пригнічуючого впливу СО2
на нейрони дихального центру.
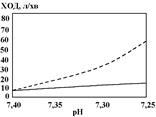 При зниженні рН артеріальної крові нижче 7,4 вентиляція легень (ХОД) також підвищується. Зменшення рН на 0,1 призводить до збільшення ХОД на 2 л/хв. Гіпервентиляція при збільшенні концентрації йонів Н+
супроводжується зниженням Рсо2
в крові. Тому, якщо дослідження проводити при постійному Рсо2
в артеріальній крові (40 мм.рт.ст.), гіпервентиляція виражена більше (заштрихована лінія).
При зниженні рН артеріальної крові нижче 7,4 вентиляція легень (ХОД) також підвищується. Зменшення рН на 0,1 призводить до збільшення ХОД на 2 л/хв. Гіпервентиляція при збільшенні концентрації йонів Н+
супроводжується зниженням Рсо2
в крові. Тому, якщо дослідження проводити при постійному Рсо2
в артеріальній крові (40 мм.рт.ст.), гіпервентиляція виражена більше (заштрихована лінія).
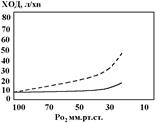 Зниження Ро2
в артеріальній крові також супроводжується гіпервентиляцією. Звертає на себе увагу той факт, що вплив зниження Ро2
впливає на ХОД, якщо величина показника стає нижчою за 50 – 60 мм.рт.ст. (норма – 100 мм.рт.ст.). тобто, гіпервентиляція розвивається у відповідь на значне зниження Ро2
і ступінь її невелика. Як і в випадку з рН, реакція стає більш вираженою, якщо при дослідженні стабілізувати Рсо2
на рівні 40 мм.рт.ст. (заштрихована лінія).
Зниження Ро2
в артеріальній крові також супроводжується гіпервентиляцією. Звертає на себе увагу той факт, що вплив зниження Ро2
впливає на ХОД, якщо величина показника стає нижчою за 50 – 60 мм.рт.ст. (норма – 100 мм.рт.ст.). тобто, гіпервентиляція розвивається у відповідь на значне зниження Ро2
і ступінь її невелика. Як і в випадку з рН, реакція стає більш вираженою, якщо при дослідженні стабілізувати Рсо2
на рівні 40 мм.рт.ст. (заштрихована лінія).
Таким чином, підвищення Рсо2 призводить до гіпервентиляції відразу ж (тобто, при збільшенні показника на 1 – 2 мм.рт.ст.) і ступінь гіпервентиляції є значною. Гіпоксія стимулює зовнішнє дихання тільки при значному зниженні Ро2 (до 50 – 60 мм.рт.ст.) і ступінь гіпервентиляції при цьому не значна.
Тому головним стимулятором зовнішнього дихання є підвищення Рсо2 в артеріальній крові, а зниження Ро2 в фізіологічних умовах рідко виступає як стимулятор зовнішнього дихання (наприклад, при підйомі в гори), та часто стає ним в умовах патології.
14. Регуляція зовнішнього дихання при фізичному навантаженні.
При фізичному навантаженні розвивається гіпервентиляція, ступінь якої пропорційна інтенсивності навантаження. Регуляція зовнішнього дихання в цих умовах здійснюється як по збуренню (ведучий механізм) так і по відхиленню (в умовах дуже інтенсивного або довготривалого навантаження).
Головним механізмом розвитку гіпервентиляції при фізичному навантаженні є безумовні рефлекси з пропріорецепторів працюючих м’язів (керуючий пристрій (КП), яким є дихальний центр, отримує по каналу зовнішнього зв’язку інформацію від пропріорецепторів працюючих м’язів про роботу що виконується; КП аналізує цю інформацію і викликає підвищення глибини та частоти дихання для того, щоб при збільшених метаболічних потребах тканин склад альвеолярного повітря залишався незмінним).
Рецептори з пропріорецепторів працюючих м’язів є ведучим, та не єдиним механізмом, який стимулює розвиток гіпервентиляції при фізичному навантаженні. Ще до початку роботи гіпервентиляція може розвиватися за механізмом умовних рефлексів (передстартові стани у спортсменів).
Від моторної зони кори ГМ паралельно з передачею команди про початок роботи до скелетних м’язів, інформація надходить і до нейронів дихального центру, що також сприяє розвитку гіпервентиляції.
Гіпервентиляція, яка розвивається внаслідок вмикання всіх вищеназваних механізмів, рідко точно відповідає метаболічним потребам організму, частіше вона буває дещо надмірною або недостатньою (дихальному центру “важко” точно оцінити сигнали, які до нього надходять з точки зору інтенсивності м’язевої роботи, а значить і метаболізму). Якщо рівень вентиляції не зовсім відповідає рівню метаболізма, проходить зміна газового складу артеріальної крові. Ці відхилення сприймаються хеморецепторами (центральними та периферичними – СП1 ) Ù передача інформації в КП каналом зворотнього зв’язку зміна вентиляції; вона стає відповідати рівню метаболізма Ù нормалізація газового складу артеріальної крові. Таким чином, хеморецептори виступають як детектори похибки і вносять необхідні корективи (рефлекторно) в роботу дихального центру при фізичному навантаженні.
І лише при роботі дуже інтенсивній чи довготривалій, коли система зовнішнього дихання не може забезпечити підтримання постійності газового складу в альвеолярному повітрі (і в артеріальній крові), рефлекси з хеморецепторів виступають як механізми стимуляції зовнішнього дихання.
15. Методи визначення енерговитрат людини. Дихальний коефіцієнт. Джерела і шляхи використання енергії в організмі людини.
З точки зору термодинаміки, організм людини є відкритою термодинамічною системою, тобто він обмінюється з навколишнім середовищем речовиною, енергією та інформацією.
Як будь-яка термодинамічна система, організм людини підкоряється законам термодинаміки:
1-й закон: загальна енергія системи та навколишнього середовища постійна, та в ході хімічних реакцій та фізичних процесів енергія може переходити із однієї форми в іншу. Тобто, 1-й закон термодинаміки представляє собою закон збереження енергії.
2-й закон: всі самовільні процеси намагаються проходити в напрямку зростання ентропії системи та навколишнього середовища. Ентропія – міра невпорядкованості системи, міра деструкції та розсіяності енергії. Тобто, 2-й закон обмежує можливі самовільні перетворення енергії в системі.
У відповідності з другим законом термодинаміки всі самовільні процеси в організмі проходять в напрямку підвищення ентропії, тобто зменшення ступеня впорядкованості структур, які складають організм. Та організм людини, як і будь-який інший організм характеризується високою ступінню впорядкованості структур. Підтримання такої впорядкованості структур можливе тільки завдяки постійному обміну енергією з зовнішнім середовищем.
Надходження енергії в організм людини проходить у вигляді енергії хімічних зв’язків харчових продуктів (жирів, білків та вуглеводів).
Перетворення енергії в організмі людини полягає в її:
- вивільненні;
- акумуляції;
- використанні.
Виділення енергії з організму людини проходить у вигляді тепла (за умові, що людини не виконує зовнішньої механічної роботи, бо при її виконанні частина енергії виділяється із організму у вигляді тепла, а частина іде на виконання роботи).
Перетворення енергії в організмі людини:
1. Вивільнення енергії.
2. Акумуляція енергії.
3. Використання енергії.
Виділення енергії з організму людини відбувається у вигляді тепла (за умови що людина не виконує жодної зовнішньої механічної роботи – тоді частина енергії виділяється у вигляді тепла, а частина йде на виконання роботи).
Перетворення енергії в організмі людини.
1. Вивільнення енергії поживних речовин відбувається при руйнуванні їх хімічних зв’язків. В організмі людини основну роль в процесах вивільнення енергїі відіграють процеси аеробного окисного фосфорилювання. Пари електронів, що вивільняються при окисненні субстратів циклу Кребса, при окисненні жирних кислот передаються на ансамбль ферментів, що називається дихальним ланцюгом. При транспорті електронів по дихальному ланцюгу зменшується їх вільна енергія. При цьому:
- частина енергії перетворюється на тепло (первинне) і виділяється з організму; ця частина складає 50-60% від усієї енергії окиснення;
- решта енергії йде на синтез АТФ з АДФ та неорганічного фосфату.
Ступінь спряження окиснення та фосфорилювання в дихальному ланцюзі характеризує відношення Фн/О2 , максимальна величина якого теоретично може досягати 3 – при витраті одного атома кисню максимально може утворитися 3 молекули АТФ і витратитися 3 атоми неорганічного фосфору. Ступінь спряження окиснення і фосфорилювання може змінюватися під впливом гормонів (катехоламінів, тироксину), при активізації симпатичного відділу вегетативної нервової системи. Вони розділяють окиснення й фосфорилювання в дихальному ланцюзі Ù більша частина енергії перетворюється на первинне тепло і менше її йде на синтез АТФ. Якщо енарговитрати організму при цьому не змінилися (для підтримки життєдіяльності та виконання роботи потрібно стільки ж енергії, як і до розділення окиснення й фосфорилювання), інтенсивність аеробного окиснення буде зростати (для синтезу такої ж кількості АТФ потрібно окиснити більше поживних речовин).
2. Акумуляція енергії відбувається у вигляді енергії макроергічних зв’язків АТФ, ГТФ, ЦТФ, КФ.
3. Використання енергії. Незалежно від умов існування організму можна виділити 3 напрямки, за котрими відбувається використання енергії макроергічних сполук:
- процеси самовідновлення; ККД цих процесів складає 25-35% (без врахування ККД синтезу АТФ, котрий складає близько 50%), решта енергії макроергів перетворюється на вторинне тепло й виділяється з організму.
- робота механізмів активного транспорту проти градієнтів концентрацій – натрій-калієва помпа зовнішньої клітинної мембрани, кальцієва помпа СПР, всмоктування в кишківнику, реабсорбція та секреція в ниркових канальцях. ККД – близько 20%.
- механічна робота скелетних та гладких м’язів.
Отже, за відсутності виконання зовнішньої роботи, вся ернергія, що використовується організмом, перетворюється на тепло і виділяється з організму. Процеси біосинтезу речовин спряжені з процесами їх розпаду, котрі йдуть з вивільненням енергії у вигляді тепла; йонні помпи створюють градієнти концентрації, за якими надалі йони рухаються пасивно, що супроводжується виділенням енергії у вигляді тепла; робота серця йде на подолання опору судин Ù виділення енергії у вигляді тепла.
Тому, виділення тепла з організму проходить у вигляді тепла (первинного та вторинного).
Методи визначення енерговитрат організму:
1. Пряма калориметрія . Дослідження проводять за допомогою спеціальних пристроїв – калориметрів, які не допускають втрати тепла в зовнішнє середовище (а саме в цьому й полягає технічна складність методу). Прилад має подвійні стінки, між якими по системі посудин рухається рідина. Організм, який знаходиться в калориметрі, виділяє тепло Ù рідина в трубах нагрівається. Розрахунок енерговитрат проводять за формулою:
![]() , де
, де
Q – тепловтрати організму;
m –маса рідини в трубах;
с – питома теплоємність рідини;
t1
– t2
– різниця температури
8-09-2015, 23:15