Устройство бактериородопсина намного проще системы хлорофилльного фотосинтеза. Белковая часть бактериородопсина представляет собой одну полипептидную цепь средней длины, которая не содержит других коферментов и простетических групп, кроме ретиналя. Бактериородопсин чрезвычайно устойчив: без потери активности его можно кипятить в автоклаве при + 130°С, изменять содержание NaCl в омывающем мембрану растворе от нуля до насыщения, в широких пределах менять рН этого раствора. Более того, можно удалить выступающие из мембраны концевые участки полипептидной цепи и даже расщепить эту цепь в одном месте по середине без ущерба для активности насоса. В то же время эффективность бактериородопсина как преобразователя энергии сравнительно низка: всего 20% энергии светового кванта превращается в ∆![]() Н
+
. При этом на один поглощенный квант через мембрану переносится один ион Н+
.
Н
+
. При этом на один поглощенный квант через мембрану переносится один ион Н+
.
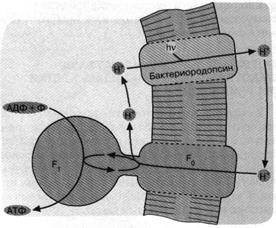
Рис. 4. Бактериородопсиновый фотосинтез со-лелюбивых архебактерий. Ионы 1-Г откачиваются из клетки бактериородопсином - белком, содержащим ретиналь в качестве хромофора, то есть группировки, поглощающей видимый свет. Ионы Н* возвращаются в клетку, двигаясь "под гору" через Н+ -АТФазный комплекс F0 F,. При этом оказывается, что Н+ - АТФаза катализирует обратную реакцию, то есть синтез АТФ, а не его гидролиз
Хлорофилльный фотосинтез
Хлорофилльный фотосинтез отличается от бактериородопсинового большей эффективностью использования светового кванта. Он устроен таким образом, что либо на каждый квант переносится через мембрану не один, а два иона Н+ , либо помимо транспорта Н+ происходит запасание энергии в форме углеводов, синтезируемых из С02 и Н2 О. Вот почему бактериородопсиновый фотосинтез был оттеснен эволюцией с авансцены. Он сохранился только у бактерий, живущих в экстремальных условиях, где более сложный и менее устойчивый хлорофилльный фотосинтез, по-видимому, просто не может существовать.
Хлорофилльный фотосинтез катализируется ферментной системой, включающей несколько белков. Квант света поглощается хлорофиллом, молекула которого, перейдя в возбужденное состояние, передает один из своих электронов в фотосинтетическую цепь переноса электронов. Эта цепь представляет собой последовательность окислительно-восстановительных ферментов и коферментов, находящихся во внутренней мембране бактерий или хлоропластов растений, где локализованы также белки, связанные с хлорофиллом. Компоненты цепи содержат, как правило, ионы металлов с переменной валентностью (железо, медь, реже марганец или никель). При этом железо может входить в состав тема (в таком случае белки называются цитохромами). Большую роль играют также негемовые железопротеиды, где ион железа связан с белком через серу цистеина или реже азот гистидина. Помимо ионов металлов роль переносчиков электронов играют производные хинонов, такие, как убихинон, пластохинон и витамины группы К.
Перенос по цепи электрона, отнятого от возбужденного хлорофилла, завершается по-разному в зависимости от типа фотосинтеза. У зеленых бактерий, использующих комплекс хлорофилла и белка, называемый фотосистемой 1 (рис. 5, а), продуктом оказывается НАДН, то есть восстановленная форма НАД+ . Восстанавливаясь, то есть присоединяя два электрона, НАД+ связывает также один Н+ . В дальнейшем образованный таким образом НАДН окисляется, передавая свой водород на различные субстраты биосинтезов.
Что касается хлорофилла, окисленного цепью, то у зеленых серных бактерий он получит недостающий электрон от сероводорода (H,S). В результате образуются также элементарная сера и ион Н+
. Белок, окисляющий H2
S, расположен на внешней поверхности бактериальной мембраны, а белок, восстанавливающий НАД+
, — на внутренней ее поверхности. Вот почему оказывается, что запускаемый светом перенос электронов от H,S к НАД"1
" образует ионы Н+
снаружи и потребляет их внутри бактерии. При этом внутренний объем клетки заряжается отрицательно относительно внешнего. Тем самым создается ∆![]() Н
+
, которая потребляется Н+
- АТФ-синтазой (комплексом факторов F0
и F,), образующей АТФ при переносе ионов Н "под гору", то есть снаружи внутрь.
Н
+
, которая потребляется Н+
- АТФ-синтазой (комплексом факторов F0
и F,), образующей АТФ при переносе ионов Н "под гору", то есть снаружи внутрь.
Другой тип бактериального фотосинтеза обнаружен у пурпурных бактерий (рис. 5. б). Здесь действует набор ферментов, отличающихся от ферментного комплекса зеленых бактерий. Это несущая хлорофилл фотосистема 2 и комплекс III. Как и в предыдущем случае, процесс начинается с поглощения кванта хлорофиллом. Первоначатьно перенос электронов происходит по фотосистеме 2.
Затем вступает комплекс III, способный транспортировать электроны сопряженно с откачкой ионов Н+ из бактерии. Процесс завершается возвращением электрона с комплекса III на хлорофилл. Что касается ионов Н+ , то они возвращаются в клетку через Н+ -АТФ-синтазу, образуя АТФ.
Отличительная черта фотосинтеза у пурпурных бактерий состоит в том, что система не нуждается во внешнем доноре электронов. Откачка ионов Н+ осуществляется путем циклического переноса электронов, поддерживаемого энергией света. Данное обстоятельство можно отнести, по-видимому, на счет эволюционного усовершенствования фотосинтеза пурпурными бактериями, которые по многим признакам являются эволюционно более продвинутой группой, чем зеленые серные бактерии.
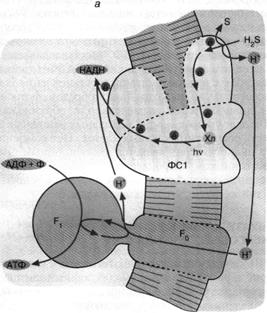
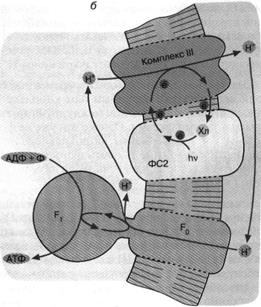
Рис. 5. Хлорофилльный фотосинтез зеленых серных (а) и пурпурных (б) бактерий: а - хлорофилл, связанный с особым белковым комплексом - фотосистемой 1 (ФС1), возбуждается квантом света и отдает электрон по цепи электронных переносчиков на НАД+ . Восстанавливаясь, НАД* связывает внутриклеточный ион НГ. Потеря электрона на хлорофилле компенсируется окислением сероводорода до серы и иона ИГ снаружи бактериальной клетки. Движение Н+ внутрь клетки через комплекс F0 F1 дает АТФ; б - хлорофилл, связанный с белком фотосистемы 2 (ФС2), поглощает квант света и запускает циклический перенос электронов. В этом процессе участвуют переносчики электронов ФС2 и дополнительного белкового комплекса III. Перенос электронов комплексом III сопряжен с откачкой ионов Н+ из клетки. Откачанные ионы I-Г возвращаются через комплекс F0 F, с образованием АТФ
Следующим шагом в эволюции фотосинтеза стали, по-видимому, цианобактерии. Цепь переноса электронов в этом случае представляет собой комбинацию: а) фотосистемы 1 зеленых бактерий, б) фотосистемы 2 и комплекса III пурпурных бактерий и в) дополнительного комплекса, расщепляющего воду на О2 и Н+ (рис. 6). Фактически донором электронов вместо сероводорода (встречающегося в достаточных количествах лишь в некоторых приданных нишах) служит вездесущая вода, запасы которой практически неограниченны. В результате конечный акцептор электоров — НАДФ+ восстанавливается, а вода окисляется. Образующийся НАДФН окисляется затем сложной системой восстановления углекислого газа до глюкозы. Таким образом, фотосинтез цианобактерии параллельно с образованием АТФ дает углевод — одно из главных резервных веществ современных живых клеток. Нет сомнений, что цианобактерия является эволюционным предшественником хлоропластов — органелл зеленыхрастений, энергетика которых устроена в основном по той же схеме, что показана на рис. 6.
ДЫХАТЕЛЬНЫЙ МЕХАНИЗМ ЭНЕРГООБЕСПЕЧЕНИЯ
Побочным продуктом фотосинтеза у цианобактерий и растений служит молекулярный кислород. Нарастание его концентрации в атмосфере привело к появлению ферментов, убирающих этот сильный окислитель, опасный для жизнедеятельности. Вероятно, первой функцией ферментов, восстанавливающих О2 до Н2 О, было снижение внутриклеточной концентрации кислорода. Однако в дальнейшем аэробная клетка научилась извлекать пользу из этого процесса, создав дыхательную цепь электронного транспорта, сопряженного с откачкой ионов Н+ .
Дыхательная цепь некоторых современных бактерий включает уже знакомый нам комплекс III, служащий связующим звеном между двумя другими белковыми комплексами. Однако это уже не фотосистемы 1 и 2, а ферменты, выполняющие функции, противоположные таковым фотосистем 1 и 2. Ферменты, о которых идет речь, были названы комплекс I и комплекс IV.
Комплекс I не восстанавливает никотинамидный нуклеотид, а окисляет его. Комплекс IV не окисляет воду до О2 , а восстанавливает О2 до воды. В итоге мы имеем сложную цепь реакций, начинающихся с окисления НАДН и кончающихся восстановлением О2 . Все три комплекса дыхательной цепи способны откачивать из клетки ионы Н+ сопряжено с переносом электронов (рис. 7).
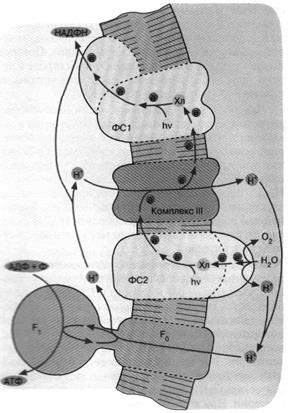
Рис. 6. Хлорофилльный фотосинтез цианобактерии. Квант света, поглощенный хлорофиллом фотосистемы 1, возбуждает перенос электронов по цепи, что завершается восстановлением НАДФ+ до НАДФН. Окисленный хлорофилл фотосистемы 1 восстанавливается комплексом III, который, в свою очередь, получает электрон от фотосистемы 2. Донирование электрона фотосистемой 2 требует еще одного кванта света (поглощаемого хлорофиллом этой фотосистемы). Потеря электрона на хлорофилле фотосистемы 2 компенсируется за счет окисления молекулы воды до О2 и Н+ . Ферменты, катализирующие всю цепь реакций переноса электрона от Н2 О до НАДФ*, расположены в мембране таким образом, что ионы Н+ откачиваются из бактериальной клетки, чтобы затем войти внутрь через FC F. и сделать АТФ. В хлоро-пластах зеленых растений происходят те же события, но ориентация всех ферментов противоположна той, которая имеет место у цианобактерии и показана на рис. 6. Соответственно у хлоропластов фотосинтетическая цепь накачивает ионы Н+ внутрь, а комплекс F0 F, переносит их наружу
Подобно тому как хлоропласты произошли от цианобактерии, митохондрии животных, растений и грибов ведут свое происхождение от аэробных бактерий. Поэтому неудивительно, что митохондриальная дыхательная цепь описывается той же схемой, что изображена на рис. 7.
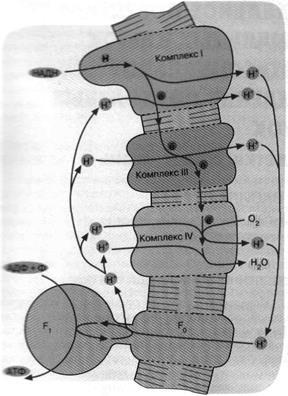
Рис.7. Механизм дыхательного фосфорилирования в аэробных бактериях и митохондриях. Ферментные комплексы I, III и IV катализируют перенос электронов от НАДН к О2 с образованием воды. Перенос электронов сопряжен с откачкой ионов Н*. Возвращение ионов Н+ через F0 F1 приводит к синтезу АТФ
ФОТОСИНТЕЗ И БИОСФЕРА
Основным и практически неиссякаемым источником энергии на поверхности Земли является энергия солнечного излучения, постоянным потоком поступающая из космоса благодаря протеканию термоядерных реакций на ближайшем к нам светиле — Солнце. Как показано на рис. 1, спектр поступающего на Землю солнечного излучения соответствует спектру излучения абсолютно черного тела, нагретого до 5900 К. Полный поток солнечного излучения (измеренный за пределами земной атмосферы), приходящийся на единицу поверхности, нормальной к направлению на Солнце, близок к 1400 Вт/м2 . Значительная часть этой энергии приходится на область видимого и ближнего инфракрасного излучения (0,3 - 1,0 мкм) — фотосинтетически активную радиацию, эффективно поглощаемую пигментами, участвующими в фотосинтезе растений и фотосинтезирующих бактерий.
Какая бы часть спектра этого излучения ни поглощалась на Земле, это в конечном счете приводит главным образом к нагреванию поверхности планеты и ее атмосферы, или же энергия вновь испускается в космическое пространство. Какова же роль фотосинтеза, фотосинтезирующих организмов в улавливании этой энергии? Почему утверждают, что фотосинтез - это энергетическая основа биологических процессов, энергетический движитель развития биосферы? Почему говорят как о фотоавтотрофии (то есть о питании за счет света) биосферы в целом, так и о фотоавтотрофии человечества, а жизнь на Земле называют космическим явлением прежде всего потому, что она существует и развивается за счет энергии, поступающей к нам из космоса — от ближайшего космического светила?
Как известно, фотосинтез растений заключается в преобразовании и запасании солнечной энергии, в результате которого из простых веществ — углекислоты и воды — синтезируются углеводы и выделяется молекулярный кислород. В общем виде этот процесс можно описать следующим уравнением (рис. 2).
Несмотря на кажущуюся простоту фотосинтеза, на Земле, пожалуй, нет более удивительного процесса, который смог бы в такой степени преобразовать нашу планету.
ЗАПАСАНИЕ ЭНЕРГИИ
Как следует из уравнения (рис. 2), на каждый ассимилированный в процессе фотосинтеза моль углекислоты запасается 114 ккал энергии. В чем же состоит достоинство запасания солнечной энергии растениями по сравнению с неорганизованной ("нефотосинтезирующей") системой? Любое вещество, поглощая квант солнечной энергии, переходит в возбужденное состояние, что уже можно рассматривать как преобразование энергии электромагнитного излучения и ее запасание. Однако энергия электронного возбуждения очень быстро (за 10-13 – 10-11 сек) растрачивается на тепло или же вновь излучается в пространство (для сложных органических молекул типа хлорофилла этот процесс происходит за 10-8 – 10-9 сек) и, следовательно, в виде возбужденных состояний энергия света может быть запасена лишь на незначительные доли секунды. В результате же фотосинтеза энергия поглощенного кванта света (или, лучше сказать, часть этой энергии) запасается надолго: от минут и часов до сотен и даже миллионов лет (как это имело место, например, при образовании горючих ископаемых — нефти, природного газа, каменного угля, торфа в результате разложения наземных и морских растений или животных). Но этим, конечно, не исчерпывается специфика фотосинтеза в использовании солнечной энергии. Так, формирование горных ледников и озер тоже происходит за счет энергии Солнца, идущей на испарение воды, и при этом тоже происходит запасание солнечной энергии на длительное время. В связи с этим говорят об еще одном преимуществе фотосинтеза: запасание солнечной энергии происходит в очень удобной для биологического использования форме - молекулярной, в виде богатых энергией связей, в основном в сахарах и их производных, а также в аминокислотах, белках, жирах, которые в любой необходимый момент могут быть использованы растениями или "съевшими" их нефотосинтезирующими (гетеротрофными) организмами для покрытия своих энергетических потребностей, для биосинтеза собственных высокомолекулярных соединений.
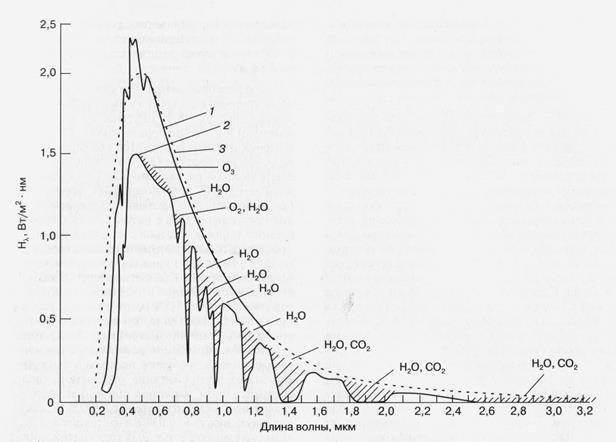
Рис. 1. Интенсивность падающего на Землю солнечного излучения (Нl ) в зависимости от длины волны. Заштрихованные области соответствуют ненаблюдаемым на уровне моря участкам спектра из-за их поглощения указанными компонентами атмосферы. 1 - Солнечное излучение за границей атмосферы, 2 - солнечное излучение на уровне моря, 3 - излучение абсолютно черного тела при 5900 К. (Справочник по геофизике и космическому пространству. Под ред. С.Л. Валлея и Мак Гроу-Хилла, Нью-Йорк, 1965).
Фотосинтез
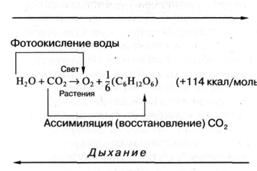
Рис. 2. Уравнение фотосинтеза кислородвыделяющих фотосинтезирующих организмов.
Масштабы фотосинтетического преобразования и запасания солнечной энергии огромны: каждый год за счет фотосинтеза на Земле образуется около 200 млрд. тонн биомассы, что эквивалентно энергии, равной 3 • 1021 Дж или 7,2 • 1020 кал. При этом необходимо иметь в виду, что фотосинтез — единственный биологический процесс, протекающий с запасанием (с увеличением) свободной энергии. Все остальные процессы, как в растениях, так и в животных, проходят за счет химической энергии, накапливаемой в фотосинтезирующих организмах в результате преобразования поглощенного солнечного света. Следовательно, практически вся живая материя на Земле представляет собой прямой или отдаленный результат фотосинтетической деятельности растений, которые являются посредниками между неиссякаемым источником энергии — Солнцем и всем живым миром нашей планеты. Именно поэтому мы говорим о фотоавтотрофии биосферы Земли, в том числе и о фотоавтотрофии человечества. Население Земли ежегодно потребляет около 1 млрд. тонн продуктов питания, что соответствует 15 • 1018 Дж, если считать численность населения равной 5 млрд. человек. Следовательно, человечество потребляет в виде органических веществ лишь около 0,5% всей энергии, запасаемой в результате фотосинтеза. Общее потребление энергии в мировом масштабе составляет 3 — 4 • 1020 Дж в год, то есть около 10% всей энергии, запасаемой за год благодаря фотосинтезу. Разведанные запасы ископаемого топлива (нефти, газа, угля, торфа) по запасенной в них энергии соответствуют продукции фотосинтетической деятельности на Земле приблизительно за 100 лет, что эквивалентно также энергии, которая содержится во всей биомассе, находящейся в настоящее время на нашей планете.
АССИМИЛЯЦИЯ ДВУОКИСИ УГЛЕРОДА
Ежегодная ассимиляция углекислого газа на Земле в результате фотосинтеза составляет около 260 млрд. тонн, что эквивалентно 7,8 • 10'° тонн углерода, и это связывание углерода компенсируется выделением практически такого же количества СО2 в результате дыхания нефотосинтезирующих организмов. Количество СО2 , вовлекаемого в цикл "фотосинтез—дыхание", составляет около 10% массы углекислого газа в атмосфере, которая в 1980 году была эквивалентна 7,1 • 10" тонн углерода. В то же время до 1860 года атмосфера содержала лишь 6,1 • 10й тонн углерода в виде СО2 , и это 15%-ное увеличение СО2 в атмосфере связывают прежде всего с появлением дополнительного источника СО2 вследствие интенсивного сжигания ископаемого топлива, которое эквивалентно в настоящее время 5 • 109 тонн углерода в год и увеличивается в среднем на 4,3% в год.
Необходимо отметить, что значительно большая часть углерода содержится в виде карбонатов в осадочных породах - 5,5 • 1016
тонн, в живых (в основном леса) и отмерших организмах - 3,5 • 1012
тонн. В мировом океане содержится в 60 раз больше углерода, чем в атмосфере (3,5 • 1013
тонн), что связано с очень высокой растворимостью СО2
в воде и образованием Н2
СО3
и, следовательно, можно было бы предположить, что незначительное дополнительное поступление СО2
в результате сжигания ископаемого топлива, которое составляет менее одного процента в год от содержания углекислого газа в атмосфере, не должно приводить к заметному увеличению содержания СО2
в атмосфере. Однако в действительности лишь в верхних слоях океана, содержащих лишь 1,5% всего углерода, растворенного в воде, обмен углерода с атмосферой осуществляется достаточно быстро (за 6 - 7 лет), тогда как для установления такого равновесия с глубинными слоями океана требуется несколько тысячелетий. Вследствие этого сжигание ископаемого топлива в промышленном масштабе привело к увеличению содержания СО2
в атмосфере с 0,027% (в доиндустриальную эпоху) до 0,034% в настоящее время. Расчеты показывают, что к 2035 году содержание углекислого газа в атмосфере удвоится, то есть будет составлять около 0,06%. Основным последствием этого, как считается, будет глобальное потепление климата, обусловленное так называемым "тепличным эффектом", связанным с тем, что углекислый
29-04-2015, 05:18