ВЫДЕЛЕНИЕ МОЛЕКУЛЯРНОГО КИСЛОРОДА
Приобретенная в процессе эволюции (более 2 млрд. лет назад) способность фотосинтезирующих растений к выделению молекулярного кислорода в результате окисления воды (см. рис. 2) привела к поистине революционным преобразованиям на Земле, из которых необходимо отметить следующие.
1. Фотосинтезирующие организмы, а через их посредство и вся живая природа, получили доступ к практически неиссякаемому и возобновляемому источнику электронов, участвующих во всех биоэнергетических процессах, — воде, что, естественно, привело к резкому возрастанию масштабов фотосинтеза и поступления энергии в биосферу. Ранее фотоокислению могли подвергаться лишь восстановленные вещества типа H2 S, представленные на Земле в довольно ограниченном количестве.
2. Продукт фотосинтетического окисления воды — молекулярный кислород, содержание которого в атмосфере возросло в результате фотосинтеза практически от нуля в древней атмосфере до 21% в настоящее время, вызвал значительные изменения во всей живой природе. Появление в атмосфере свободного О2 привело к массовой гибели существовавших тогда организмов, для которых такой сильный окислитель как кислород оказался сильнейшим ядом, поскольку он резко нарушал протекание биоэнергетических процессов. В то же время в результате длительной эволюции на фоне возрастающей концентрации О2 в атмосфере появились новые, аэробные организмы, обратившие наличие О2 себе на пользу. Они сумели включить его в свой метаболизм в качестве эффективного конечного акцептора электронов в дыхательной цепи. Это позволило поднять биоэнергетику на новый, значительно более высокий уровень, так как в этом случае происходит более эффективное "сжигание" органики. Так, например, выход макроэргического эквивалента — аденозинтрифосфата (АТФ) на одну молекулу исходной гексозы составляет две молекулы при молочнокислом брожении и 36 — 38 молекул (то есть в 18—19 раз выше) при аэробном дыхании.
Фотосинтетическое образование О2 , сопровождающееся поглощением СО2 , в сочетании с появившейся возможностью аэробного дыхания, при котором происходит обратный процесс — потребление 02 , сопровождающееся освобождением СО2 , привело к образованию замкнутых циклов О2 и СО2 , имеющих глобальные масштабы. Сочетание этих циклов позволяет восполнять содержание как СО2 , так и О2 , а их участие в едином цикле "фотосинтез-дыхание" обеспечивает постоянное поступление энергии в биосферу (см. рис. 2).
3. Появление кислородвыделяющих организмов привело к тому, что практически все процессы на поверхности Земли приняли биогеохимический характер. Именно в этот период значительного повышения О2 в атмосфере произошло окисление соединений железа, серы, марганца, причем на это ушло более 95% кислорода, выделенного фотосинтезирующими организмами за всю историю биосферы. Атмосфера Земли до появления кислородвыделяющих фотосинтезирующих организмов была разреженной и состояла главным образом из СО2 и аммиака. По мере возрастания О2 в атмосфере значительно увеличивается и содержание молекулярного азота благодаря окислению аммиака. Сам процесс фотосинтеза, сопровождающийся образованием и запасанием органического вещества, привел к значительному, более чем в 100 раз, уменьшению содержания СО2 в атмосфере.
В результате первичная разреженная атмосфера превратилась во вторичную плотную азотокислородную оболочку Земли. Теперь солнечная и другие виды космической радиации, прежде чем достичь поверхности, должны были фильтроваться через мощный слой атмосферы. Кроме того, под действием фотохимического процесса в верхних слоях атмосферы двухатомный молекулярный кислород превращается в трехатомную молекулу озона, имеющего интенсивное поглощение в области жесткого ультрафиолета (0,2 — 0,3 мкм). В результате образовавшийся озоновый экран стал задерживать наиболее опасную для живых организмов ультрафиолетовую радиацию.
Таким образом, фотосинтез, основанный на окислении воды, благодаря возможности использовать воду в качестве субстрата стал мощным поставщиком энергии в биосферу, революционным образом обогатил биоэнергетику благодаря включению О2 в метаболизм живых организмов, защитил живую материю от гибельного действия космической радиации. Все это индуцировало мощное развитие биологической материи на Земле, приведшее в конечном счете к появлению человека. И в настоящее время фотосинтез лежит в основе биологического круговорота энергии и веществ на Земле, от масштабов которого зависит и жизнь на нашей планете, и ее разнообразие и возможности.
МОЛЕКУЛЯРНЫЕ ОСНОВЫ ПРЕОБРАЗОВАНИЯ И ЗАПАСАНИЯ ЭНЕРГИИ СВЕТА ПРИ ФОТОСИНТЕЗЕ
Живая природа настолько рационально решает проблемы эффективного улавливания солнечной энергии, ее преобразования и запасания в удобной для биологического потребления форме, что до сих пор не удается воспроизвести основные этапы фотосинтеза в искусственных системах. Первичное улавливание и преобразование энергии света при фотосинтезе происходит с непременным участием хлорофилла, что было выявлено уже в классических работах К.А. Тимирязева, показавшего, что именно та часть солнечного света, которая поглощается хлорофиллом, эффективно инициирует процесс фотосинтеза.
Как показано на рис. 3а, молекула хлорофилла состоит из порфиринового ядра, образованного тетрапиррольным кольцом с расположенным в центре атомом магния, и фитольного хвоста. Хлорофилл относится к классу веществ, называемых пигментами, то есть сильноокрашенных соединений, характеризующихся интенсивным поглощением видимого света. Было установлено, что система "легко возбудимых" сопряженных связей порфиринового кольца обусловливает высокую способность хлорофилла к поглощению света (рис. 36). Так, миллиметровый слой 1 мМ раствора хлорофилла поглощает около 90% падающего на него красного или синего света (зеленый свет поглощается менее эффективно, что и определяет зеленый цвет растений). Для хлорофилла, как и для других пигментов, характерно относительно длительное время, около 1(Г8 сек, нахождения в возбужденном состоянии, что важно для участия хлорофилла в фотохимических реакциях. У многих других пигментов, например, у производных хлорофилла, у которых центральный атом магния заменен на медь или марганец, это время гораздо ниже вследствие чрезвычайно быстрого перехода энергии электронного возбуждения в тепло.
Для выяснения молекулярных основ участия хлорофилла в первичных процессах преобразования солнечной энергии при фотосинтезе важнейшее значение имело открытие, сделанное нашим соотечественником, академиком А.А. Красновским, который показал, что при переходе в возбужденное состояние в результате поглощения кванта света молекула хлорофилла приобретает свойство вступать в такие окислительно-восстановительные (то есть идущие с переносом электронов) реакции, к которым она не была способна в темноте. При этом молекула хлорофилла (Хл) может потерять свой электрон, то есть окислиться, отдав его другому веществу, называемому в данном случае акцептором (А) электрона, тогда как хлорофилл выступает в данной реакции как донор электрона. Эта реакция получила название реакции Красновского. Возбужденная молекула хлорофилла (Хл*) может также восстанавливаться, то есть принять "лишний" электрон, отобрав его у другого вещества, служащего в данном случае донором (Д) электрона. Если помимо Хл в системе присутствуют Д и А, то в результате сочетания процессов окисления Хл и его восстановления свет, поглощаемый хлорофиллом, будет индуцировать перенос электрона от Д к А с образованием Д+ (окисленного Д) и А- (восстановленного А). Хлорофилл при этом, несмотря на участие в двух ступенях переноса электрона, возвращается в исходное состояние готовности к новому фотохимическому циклу переноса электрона, согласно реакции
![]()
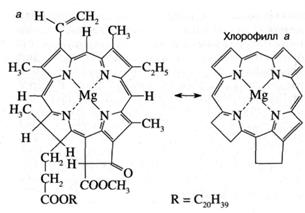
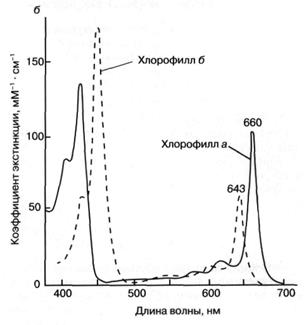
Рис. 3. Структурная химическая формула (а) и спектр поглощения (зависимость поглощения от длины волны) (б) хлорофилла.
В результате получается система с разделенны-мрядами Д+ ХлА- (элементарная электрическая баторейка), в которой может быть запасена значительная часть энергии кванта света, поглощенного хлорофиллом. Как показано в работах последних десятилетий, именно этот принцип — фоторазделениязарядов — лежит в основе первичного преобразования энергии света при фотосинтезе.
Как показано на рис. 4, хлорофилл, определяющий зеленый цвет листа растения, распределен не повсей растительной клетке, а сосредоточен в пластидах, называемых хлоропластами. В хлоропластах сосредоточен фотосинтетический аппарат, и их роль состоит в энергообеспечении клетки за счет поглощенного света. Хлоропласт имеет геном, который делится при делении хлоропласта. Несмотря на наличие собственного генома, обеспечивающего синтез большей части структур фотосинтетического аппарата, ряд строго необходимых для протекания фотосинтеза компонентов (например, основные ферменты, участвующие в биосинтезе хлорофилла, некоторые белки, входящие в систему фотосинтетического окисления воды), кодируются в клеточном геноме. Хлоропласт заполнен так называемыми тилакоидами - замкнутыми сплющенными "мешочками", которые состоят из двухслойных липидных мембран, образованных в результате впячивания внутреннего слоя двухслойной мембраны, ограничивающей весь хлоропласт от цитоплазмы. Тилакоиды сгруппированы в плотно упакованные темно-зеленые "стопки", получившие название гран, которые связаны между собой частью тилакоидов. В мембраны тилакоидов погружены белки или белковые комплексы, большая часть которых насквозь пронизывает мембрану. Только часть из этих комплексов окрашена в зеленый цвет, то есть содержит хлорофилл; было установлено, что практически весь хлорофилл находится в хлорофилл-белковых комплексах, или, лучше сказать, в пигмент-белковых комплексах, поскольку все они наряду с хлорофиллом содержат другие пигменты.
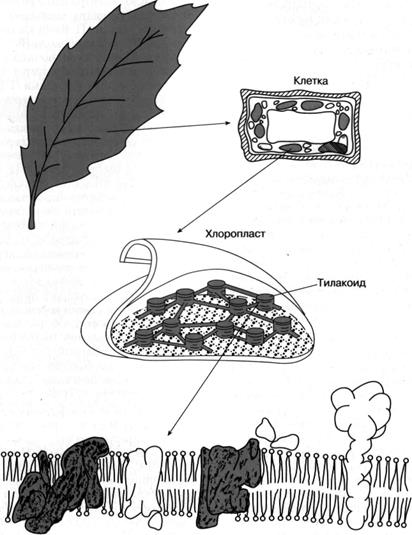
Мембрана тилакоида Рис. 4. Локализация фотосинтетического аппарата в клетке зеленого растения.
Пигмент-белковые комплексы можно разделить на две функциональные группы. Основная функция первой из них — "светособирающих" или "антенных" пигмент-белковых комплексов - состоит в эффективном улавливании энергии света. Более 90% всего хлорофилла сосредоточено именно в "антенных" комплексах. Большая их часть представляет собой белки с молекулярной массой от 20000 до 70000, на каждом из которых расположены 10 - 40 молекул хлорофилла. Помимо основного пигмента — хлорофилла а — в состав этих комплексов входит его аналог — хлорофилл б, а также каротиноиды — пигменты желтого или оранжевого цвета (один из них, бета-каротин, определяет окраску моркови). Цианобактерии содержат также бесхлорофилльные пигмент-белковые комплексы - фикобилины, окрашенные в синий или красный цвет. Они эффективно поглощают свет в области спектра, где хлорофилл имеет слабое поглощение, что позволяет осуществить более эффективное улавливание солнечной энергии.
Энергия возбуждения от светособирающих пигментов с очень высокой эффективностью (близкой к 100%) передается на хлорофилл другой группы пигмент-белковых комплексов - фотохимические реакционные центры. Их основная и важнейшая для всей биосферы функция состоит в том, чтобы преобразовать энергию электронного возбуждения хлорофилла (полученную в результате поглощения света в данном комплексе или в результате миграции возбуждения от светособирающих комплексов) в энергию разделенных зарядов, подобно тому, как это происходит в реакции Красновского, описанной выше.
У кислородвыделяющих организмов имеется два типа реакционных центров, функционирующих в так называемых фотосистеме 1 (ФС-1) и фотосистеме 2 (ФС-2) (рис. 5). Аналоги каждого из этих реакционных центров обнаружены у ряда фотосинте-зируюших бактерий, и молекулярная структура одного из них, выделенного из пурпурной бактерии Rhodopseudomonasviridis, впервые была установлена с помощью рентгеноструктурного анализа. Необходимо отметить, что значительная часть знаний о структурной и функциональной организации фотосинтетических реакционных центров получена при помощи изучения именно бактериальных реакционных центров. Они отличаются высокой стабильностью в изолированном из мембраны состоянии, и именно из них удалось впервые получить
кристаллы, использование которых в рентгено-структурных исследованиях позволило получить данные о молекулярной структуре реакционного центра. За эту работу группа немецких исследователей в 1988 году получила Нобелевскую премию.
Бактериальный реакционный центр состоит из трех полипептидов с молекулярной массой от 24000 до 32000, на которых расположены молекулы одного из представителей хлорофиллов - бактериохлорофилла (Бхл) и две молекулы его безмагниевого производного — бактериофеофитина (Бфф). Две из четырех молекул Бхл расположены настолько близко друг к другу (около 3 Å), что формируют компактное образование — димер (П), а четыре другие одиночные молекулы пигментов образуют две симметричные по отношению к П цепочки Бхл—Бфф. Энергия электронного возбуждения, полученная в результате поглощения света пигментами самого реакционного центра или в результате ее миграции от светособирающих комплексов, локализуется в конечном счете на П. Возбужденный П, обозначаемый как П*, за время около 10-12 сек передает свой электрон на одну из одиночных молекул пигмента, так что в результате образуется первичное состояние с разделенными зарядами П+ Бфф- . Это состояние обладает очень коротким временем жизни (около 10~8 сек) вследствие возможного возврата электрона от Бфео- к П+ . Однако вероятность этого процесса очень низка благодаря более быстрому (2 • 10-10 сек) "прямому" переносу от Бфео- к "стабильному" акцептору электрона, получившему название Q. При этом образуется состояние П+ БфеоQ- , время жизни которого уже составляет около 10-3 сек. Необходимо отметить очень высокую эффективность описанных стадий фоторазделения зарядов. Так, квантовая эффективность, то есть доля поглощенных квантов, вызвавших перенос электрона, близка к 100%. Энергетическая эффективность, то есть доля энергии возбуждения, запасенной в результате фотохимического акта, составляет свыше 90% для первой стадии переноса электрона и около 50 - 70% для второй. Об изумительном совершенстве этой уникальной биологической "машины" свидетельствует, например, тот факт, что с такой же эффективностью фотоперенос электрона в реакционном центре осуществляется даже при температуре жидкого гелия (-271°С).
Основным итогом фотохимической стадии преобразования энергии света у кислородвыделяющих фотосинтезируюших организмов является фотоперенос электронов от воды к НАДФ+ . Обе фотосистемы (ФС-1 и ФС-2) участвуют в этом процессе, обеспечивая двухступенчатое поступление энергии для его протекания. ФС-2 осуществляет окисление воды с образованием молекулярного кислорода, согласно реакции: 2Н2 0 + 4hv—О2 + 4е+ 4Н+ , где hvобозначает квант света, е— электрон. В результате фотохимического акта реакционного центра ФС-2 образуется самый сильный биологический окислитель — окисленный хлорофилл, который окисляет воду с участием марганецсодержащей энзиматической системы. Электроны, оторванные от воды, через цепь темновых реакций поступают на ФС-1, использующую их для фотовосстановления НАДФ+ до НАДФН, которое тоже осуществляется с участием специальной энзиматической системы. Окисление воды, а также перенос электронов от ФС-2 к ФС-1 приводит к появлению разности концент-
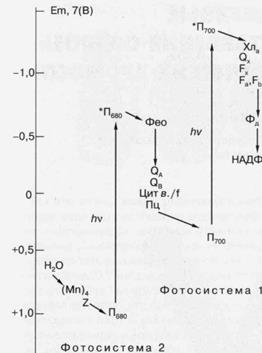
Рис. 5. Схематическое изображение фотосинтетической цепи переноса электрона в хлоропласах растений и цианобактерий. (Мп)4 - Комплекс из четырех атомов Мп, связанных с белками реакционного центра фотосистемы 2 (ФС-2); Z - вторичный донор электрона ФС-2, (остаток тирозина); Пбао - первичный донор электрона ФС-2 (димер хлорофилла); *П680 - возбужденное состояние хлорофилла П680 ; Фео - первичный акцептор электрона ФС-2, феофитин; QA и QB -акцепторы электрона хиноновой природы; цит в./ f - комплекс цитохромов, участвующих в переносе электрона от ФС-2 к фотосистеме 1 (ФС-1); Пц - пластоцианин (подвижный переносчик электрона); П700 - первичный донор электрона ФС-1 (димер хлорофилла); *П700 - возбужденное состояние П700 ;Хла -(хлорофилл) и Ох -(хинон), соответственно, первичный и вторичный акцепторы электрона ФС-1; Fx , Fa и Fb акцепторы электрона ФС-1 (Fe-S-центры); Фд - ферредоксинрастворимый переносчик электрона (Fe-S-содержащий белок); НАДФ+ - никотинамидаденин динуклеотид фосфат (конечный переносчик электронов, используемый вместе с АТФ в ассимиляции СО2 ), hv - квант света. По вертикальной шкале указаны приблизительные значения окислительно-восстановительных потенциалов переносчиков электрона при рН 7.
раций ионов Н+
по обе стороны тилакоидной мембраны, которая, как и в случае митохондрий, необходима для осуществления процесса фосфо-рилирования — образования АТФ, основного энергетического эквивалента, используемого в качестве источника энергии в биологических процессах. Образованные в результате фотохимического переноса электронов АТФ и НАДФН используются для восстановления СО2
с образованием первичных са-харов, которое в упрощенном виде можно описать следующим образом: СО2
+ 4е + 4Н+
—*•
СН2
О + + Н2
О. При этом на каждую молекулу СО2
расходуется две молекулы НАДФН и три молекулы АТФ. Этот процесс, в результате которого "восстановленная" молекула СО2
включается в состав гексозы, осуществляется через ряд стадий с участием сложного цикла энзиматических реакций, получившего название цикла Кальвина по имени его открывателя. В заключение необходимо отметить, что исследование фотосинтеза — сложнейшего фундаментального биологического процесса, имеющее давние традиции в отечественной науке, привлекает в настоящее время внимание все большего числа естествоиспытателей — биологов, физиков, химиков, математиков. Познание молекулярных механизмов фотосинтеза будет иметь большое значение для обеспечения человечества экологически чистой энергией за счет практически неиссякаемого источника — солнечного излучения (например, на основе фоторазложения воды на молекулярной водород и кислород), для повышения фотосинтетической продуктивности растений, лежащей в основе обеспечения человечества пищей, для использования принципов фотопреобразования световой
29-04-2015, 05:18