По ряду соображений именно РНК, а не ДНК, могла представлять собой первичный генетический материал.
Во-первых, и в химическом синтезе, и в биохимических реакциях рибонуклеотиды предшествуют дезоксирибонуклеотидам; дезоксирибонуклеотиды - продукты модификации рибонуклеотидов (см. рис. 2).
Во-вторых, в самых древних, универсальных процессах жизненного метаболизма широко представлены именно рибонуклеотиды, а не дезоксирибонуклеотиды, включая основные энергетические носители типа рибонуклеозид-полифосфатов (АТФ и т.п.).
В-третьих, репликация РНК может происходить без какого бы то ни было участия ДНК, а механизм редупликации ДНК даже в современном живом мире требует обязательного участия РНК-затравки в инициации синтеза цепи ДНК.
В-четвертых, обладая всеми теми же матричными и генетическими функциями, что и ДНК, РНК способна также к выполнению ряда функций, присущих белкам, включая катализ химических реакций. Таким образом, имеются все основания рассматривать ДНК как более позднее эволюционное приобретение - как модификацию РНК, специализированную для выполнения функции воспроизведения и хранения уникальных копий генов в составе клеточного генома без непосредственного участия в биосинтезе белков.
После того как были открыты каталитически активные РНК, идея первичности РНК в происхождении жизни получила сильнейший толчок к развитию, и была сформулирована концепция самодостаточного мира РНК,
предшествовавшего современной жизни [16, 17]. Возможная схема возникновения мира РНК представлена на рис. 6.
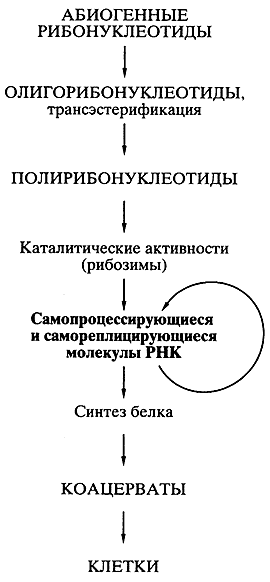 |
Рис. 6.
Схематическое представление пути происхождения жизни согласно современной концепции первичности мира РНК |
Абиогенный синтез рибонуклеотидов и их ковалентное объединение в олигомеры и полимеры типа РНК могли происходить приблизительно в тех же условиях и в той же химической обстановке, что постулировались для образования аминокислот и полипептидов. Недавно А.Б. Четверин с сотрудниками (Институт белка РАН) экспериментально показали, что по крайней мере некоторые полирибонуклеотиды (РНК) в обычной водной среде способны к спонтанной рекомбинации, то есть обмену отрезками цепи, путем транс-эстерификации [18]. Обмен коротких отрезков цепи на длинные, должен приводить к удлинению полирибонуклеотидов (РНК), а сама подобная рекомбинация способствовать структурному многообразию этих молекул. Среди них могли возникать и каталитически активные молекулы РНК.
Даже крайне редкое появление единичных молекул РНК, которые были способны катализировать полимеризацию рибонуклеотидов или соединение (сплайсинг) олигонуклеотидов на комплементарной цепи как на матрице [19, 20], означало становление механизма репликации РНК. Репликация самих РНК-катализаторов (рибозимов) должна была повлечь за собой возникновение самореплицирующихся популяций РНК. Продуцируя свои копии, РНК размножались. Неизбежные ошибки в копировании (мутации) и рекомбинации в самореплицирующихся популяциях РНК создавали все большее разнообразие этого мира. Таким образом, предполагаемый древний мир РНК - это "самодостаточный биологический мир, в котором молекулы РНК функционировали и как генетический материал, и как энзимоподобные катализаторы" [21].
Возникновение биосинтеза белка. Далее на основе мира РНК должно было происходить становление механизмов биосинтеза белка, появление разнообразных белков с наследуемой структурой и свойствами, компартментализация систем биосинтеза белка и белковых наборов, возможно, в форме коацерватов и эволюция последних в клеточные структуры - живые клетки (см. рис. 6).
Проблема перехода от древнего мира РНК к современному белок-синтезирующему миру - наиболее трудная даже для чисто теоретического решения. Возможность абиогенного синтеза по-липептидов и белковоподобных веществ не помогает в решении проблемы, так как не просматривается никакого конкретного пути, как этот синтез мог бы быть сопряжен с РНК и подпасть под генетический контроль. Генетически контролируемый синтез полипептидов и белков должен был развиваться независимо от первичного абиогенного синтеза, своим путем, на базе уже существовавшего мира РНК. В литературе предложено несколько гипотез происхождения современного механизма биосинтеза белка в мире РНК, но, пожалуй, ни одна из них не может рассматриваться как детально продуманная и безупречная с точки зрения физико-химических возможностей. Представлю свою версию процесса эволюции и специализации РНК, ведущего к возникновению аппарата биосинтеза белка (рис. 7), но и она не претендует на законченность.
Предлагаемая гипотетическая схема содержит два существенных момента, кажущихся принципиальными.
Во-первых, постулируется, что абиогенно синтезируемые олигорибонуклеотиды активно рекомбинировали посредством механизма спонтанной неэнзиматической трансэстерификации [18], приводя к образованию удлиненных цепей РНК и давая начало их многообразию. Именно этим путем в популяции олигонуклеотидов и полинуклеотидов и могли появиться как каталитически активные виды РНК (рибозимы), так и другие виды РНК со специализированными функциями (см. рис. 7). Более того, неэнзиматическая рекомбинация олигонуклеотидов, комплементарно связывающихся с полинуклеотидной матрицей, могла обеспечить сшивание (сплайсинг) фрагментов, комплементарных этой матрице, в единую цепь. Именно таким способом, а не катализируемой полимеризацией мононуклеотидов, могло осуществляться первичные копирование (размножение) РНК. Разумеется, если появлялись рибозимы, обладавшие полимеразной активностью [20], то эффективность (точность, скорость и продуктивность) копирования на комплементарной. матрице должна была значительно возрастать.
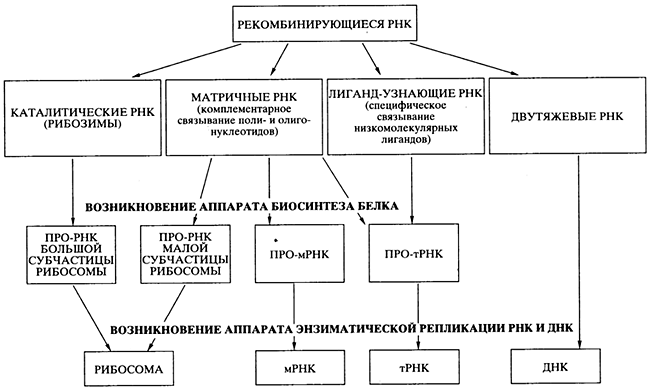
Рис. 7.
Схема эволюции и специализации молекул РНК
в процессе перехода от древнего мира РНК к современному миру
генетически детерминированного биосинтеза белков
Второй принципиальный момент в моей версии состоит в том, что первичный аппарат биосинтеза белка возник на базе нескольких видов специализированных РНК до появления аппарата энзиматической (полимеразной) репликации генетического материала - РНК и ДНК. Этот первичный аппарат включал каталитически активную прорибосомную РНК, обладавшую пептидил-трансферазной активностью; набор про-тРНК, специфически связывающих аминокислоты или короткие пептиды; другую прорибосомную РНК, способную взаимодействовать одновременно с каталитической прорибосомной РНК, про-мРНК и про-тРНК (см. рис. 7). Такая система уже могла синтезировать полипептидные цепи за счет катализируемой ею реакции транспептидации. Среди прочих каталитически активных белков - первичных ферментов (энзимов) - появились и белки, катализирующие полимеризацию нуклеотидов - репликазы, или НК-полимеразы.
Впрочем, возможно, что гипотеза о древнем мире РНК как предшественнике современного живого мира так и не сможет получить достаточного обоснования для преодоления основной трудности - научно правдоподобного описания механизма перехода от РНК и ее репликации к биосинтезу белка. Имеется привлекательная и детально продуманная альтернативная гипотеза А.Д. Альтштейна (Институт биологии гена РАН), в которой постулируется, что репликация генетического материала и его трансляция - синтез белка - возникали и эволюционировали одновременно и сопряженно, начиная с взаимодействия абиогенно синтезирующихся олигонуклеотидов и аминоацил-нуклеотидилатов - смешанных ангидридов аминокислот и нуклеотидов [22]. Но это уже следующая сказка... ("И Шахразаду застигло утро, и она прекратила дозволенные речи" .)
Литература
1. Watson J.D., Crick F.H.C. Molecular structure of nucleic acids // Nature. 1953. V. 171. P. 738-740.
2. Watson J.D., Crick F.H.C. Genetic implications of the structure of deoxyribose nucleic acid // Nature 1953 V. 171. P. 964-967.
3. Спирин А.С. Современная биология и биологическая безопасность // Вестник РАН. 1997. № 7.
4. Spirin A.S. On macromolecular structure of native high-polymer ribonucleic acid in solution // Journal of Molecular Biology. 1960. V. 2. P. 436-446.
5. Kirn S.H., Suddath F.L., Quigley GJ. et al. Three-dimensional tertiary structure of yeast phenylalanine transfer RNA // Science. 1974. V. 185. P. 435-40.
6. Robertas J.D., Ladner J.E., Finch J.T. et al. Structure of yeast phenylalanine tRNA at 3 A resolution // Nature. 1974. V. 250. P. 546-551.
7. Vasiliev V.D., Serdyuk I.N., Gudkov A.T., SPIRin A.S. Self-organization of ribosomal RNA // Sturcture, Function and Genetics of Ribosomes / Eds. Hardesty B. and Kramer G. New York: Springer-Verlag, 1986. P. 129-142.
8. Baserga SJ., Steitz J.A. The diverse world of small ribo-nucleoproteins // The RNA World / Eds. Gesteland R.F. and Atkins J.F. New York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993. P. 359-381.
9. Kruger К., Grabowski PJ., Zaug AJ. et al. Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA intervening sequence of Tetrahymena // Cell. 1982. V. 31. P. 147-157.
10. Guerrier-Takada С., Gardiner К., Marsh Т. et al. The RNA moiety of ribonucleases P is the catalytic subunit of the enzyme // Cell. 1983. V. 35. P. 849-857.
11. Опарин А.И. Происхождение жизни. М.: Московский рабочий, 1924.
12. Опарин А.И. Возникновение жизни на Земле (3-е изд.). М.: Изд-во АН СССР, 1957.
13. Woese С. The evolution of the genetic code // The Genetic Code. New York: Harper & Row, 1967. P. 179-195.
14. Crick F.H.C. The origin of the genetic code // Journal of Molecular Biology. 1968. V. 38. P. 367-379.
15. Orgel L.E. Evolution of the genetic apparatus // Journal of Molecular Biology. 1968. V. 38. P. 381-393.
16. Gilbert W. The RNA world // Nature. 1986. V 319 P. 618.
17. Joyce G.F., Orgel L.E. Prospects for understanding the origin of the RNA world // The RNA World / Eds. Gesteland R.F. and Atkins J.F. New York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993 P 1-25.
18. Chetverina H.V., Demidenko A.A., Ugarov V.I., Chetverin A.B. Spontaneous rearrangements in RNA sequences // FEBS Letters. 1999. V. 450. P. 89-94.
19. Bartel D.P., Szostak J.W. Isolation of new ribozymes from a large pool of random sequences // Science. 1993. V. 261. P. 1411-1418.
20. Ekland E.H., Bartel D.P. RNA-catalysed RNA polymerization using nucleoside triphosphates // Nature. 1996 V. 382. P. 373-376.
21. Orgel L.E. The origin of life - a review of facts and speculations //Trends in Biochemical Sciences. 1998. V. 23. p. 491-495.
22. Альтштейн А.Д. Происхождение генетической системы: гипотеза прогенов // Молекулярная биология. 1987. Т. 21. С. 309-322.
29-04-2015, 01:58