Зоны относительной мощности мышечной работы
В настоящее время приняты различные классификации мощности мышечной деятельности. Одна из них – классификация по В.С.Фарфелю, базирующаяся на положении о том, что мощность выполняемой физической нагрузки обусловлена соотношением между тремя основными путями ресинтеза АТФ, функционирующими в мышцах во время работы. Согласно этой классификации выделяют четыре зоны относительной мощности мышечной работы: максимальной, субмаксимальной, большой и умеренной.
Работа в зоне максимальной мощности может продолжаться в течение 15-20 с. Основной источник АТФ в этих условиях – креатин-фосфат. Только в конце работы креатинфосфатная реакция замещается гликолизом.
Работа в зоне субмаксимальной мощности имеет продолжительность до 5 мин. Ведущий механизм ресинтенза АТФ – гликолитический. В начале работы, пока гликолиз не достиг максимальной скорости, образование АТФ идет за счет креатинфосфата, а в конце работы гликолиз начинает заменяться тканевым дыханием. Работа в зоне субмаксимальной мощности характеризуется самым большим кислородным долгом.
При беге на длинные дистанции (5 и 10 км) аэробное окисление углеводов является основным механизмом энергообеспечения работы, так как на его долю приходится до 87 % общих затрат энергии на дистанции 5 км и около 97 % — на дистанции 10 км (рис. 136). На этих дистанциях вклад анаэробных источников энергии также еще достаточно большой. Он может достигать 15 % общих затрат энергии и играет важную роль при финишном ускорении, приносящем победу при беге на длинные дистанции. Наиболее значительным фактором, влияющим на работу на выносливость, является кислородное снабжение работающих мышц, поскольку потребление кисло- зда во время бега поддерживает максимальную скорость о^игле^ия углеводов. Порог анаэробного обмена у стайеров при работе ^ ся при 75—90 % МПК.
При марафонском беге затраты энергии восполняются исключительно за счет аэробного процесса. Погашение этих затрат невозможно только за счет окисления углеводов из-за недостаточности запасов гликогена в работающих мышцах спортсмена, поэтому значительная часть энергии обра- .ется за счет окисления жиров, на долю которых может приходиться от 10 до 50 % общих затрат энергии (рис. 137). Вклад жиров в энергообеспечение при мышечной работе на длинных и сверхдлинных дистанциях у -юокотренированных бегунов с большими запасами гликогена в работащих мышцах составляет 12—20%, у нетренированных бегунов — более 60 %. Всего на дистанции марафонского бега окисляется около 300 г жи-гов, Использование жиров в качестве источника энергии менее эффективно сравнению с окислением углеводов, так как происходит оно с более экой скоростью и с большим потреблением кислорода. Увеличивающаяся концентрация жирных кислот в крови при мобилизации жиров и высокая скорость их окисления в работающих мышцах тормозят окисление в
них углеводов, в основном глюкозы крови, что влечет за собой снижение
скорости бега. Увеличение доли окисляемых жиров при работе способ-
ствует также развитию центрального торможения. Мобилизация жиров при физической работе активизируется рядом гормонов.
При длительной работе наряду с увеличением использования в энерге-
тическом обмене жиров может происходить новообразование углеводов и?
веществ неуглеводной природы (глюконеогенез), активируемое гормонов
кортизолом. Основным субстратом глюконеогенеза являются аминокислоты
часть которых накапливается в мышцах при работе в результате распад
тканевых белков. Они могут быть использованы для образования глюкозы.
Данные об участии различных источников энергии в обеспечении ре
синтеза АТФ при беге на разные дистанции приведены в табл. 26. Вклз,
отдельных энергетических источников в обеспечение работы имеет важ
ное значение при выборе оптимальной стратегии подготовки в избранном
виде спорта, в том числе близких по интенсивности и продолжительности
к рассмотренным физическим беговым нагрузкам.
3) Описать биохимические изменения в мышцах под влиянием тренировки скоростно-силовой направленности.
В процессе мышечной деятельности изменяется скорость энергетического обмена в сердечной мышце (миокарде). Сердечная мышца пронизана густой сетью кровеносных капилляров, доставляющих большое количество кислорода, и имеет высокую активность ферментов аэробного обмена, поэтому в ней преобладают аэробные энергетические реакции. В состоянии относительного покоя основными источниками энергии для сердечной мышцы являются жирные кислоты, кетоновые тела и глюкоза, доставляемые кровью. При напряженной мышечной деятельности миокард усиленно поглощает из крови и окисляет молочную кислоту, поэтому запас гликогена в нем почти не расходуется.
В головном мозгу во время мышечной деятельности усиливается энергетический обмен, что выражается в увеличении потребления мозгом глюкозы и кислорода из крови, повышении скорости обновления гликогена и фосфолипидов, усилении распада белков и накоплении аммиака. Мозг, как и сердце, снабжается энергией за счет аэробных процессов. При работе большой мощности либо при очень продолжительной работе может снижаться запас макроэргических фосфатов в нервных клетках, что является одним из факторов развития утомления.
Биохимические изменения, происходящие в скелетных мышцах во время работы, обычно определяют по содержанию продуктов метаболизма мышц в крови, моче, выдыхаемом воздухе либо непосредственно в :ышцах (см. главу 24).
В качестве показателя интенсивности и емкости аэробных механизмов энергообеспечения часто используется величина максимального потребления кислорода. Степень вовлечения гликолиза в энергетическое обеспечение мышц можно оценить путем измерения в первые минуты восстанов- ения после работы содержания молочной кислоты в крови, а креатинфосфокиназной реакции — по содержанию в крови продуктов распада КрФ — креатина и креатинина. О включении жиров в энергетические реакции можно судить по содержанию в крови свободных жирных кислот и кетоновых тел. По показателям кислотно-щелочного равновесия можно сделать вывод о способности организма противостоять неблагоприятному действию кислотных продуктов анаэробного обмена и т. п.
Однако содержание промежуточных продуктов обмена в крови зависит т скорости их образования в клетках, диффузии через клеточные мембра- з1, а также от потребления их различными тканями. Поэтому один и тот же указатель изменения в мышечных клетках, измеренный в крови или •эчени, будет отражать изменения в тканях с разной степенью точности, ак, о скорости мобилизации углеводных запасов печени свидетельствует
содержание глюкозы в крови. В начале работы, а также пр^Пфатковремен- ной мощной работе концентрация глюкозы в крови, как правило, повышается, что свидетельствует о повышении скорости мобилизации гликогена и незначительном использовании глюкозы мышцами. При работе в условиях устойчивого состояния ее содержание в крови близко к уровню покоя, так как скорость поступления в кровь и скорость ее использования мышцами примерно одинаковы. При длительной работе концентрация глюкозы в крови может быть ниже уровня покоя, поскольку снижаются запасы гликогена печени и скорость его мобилизации, а потребность тканей в глюкозе продолжает оставаться высокой.
При интенсивной гликолитической работе в мышцах резко увеличивается содержание молочной кислоты. Она способна быстро диффундировать из работающих мышц в кровь, где ее уровень резко повышается, а окисление во время напряженной работы протекает с относительно малой скоростью, поэтому содержание молочной кислоты в крови в определенной степени отражает скорость образования ее в скелетных мышцах. В состоянии покоя концентрация молочной кислоты в крови составляет 1,1— 2,2 ммоль ■ л-1 (0,1—0,2 г • л-1 ).
При выполнении легкой и умеренно тяжелой работы (с уровнем кислородного запроса около 50 % МПК) прирост концентрации молочной кислоты в крови невелик (до 0,4—0,5 г ■ л"1 ), а при выполнении продолжительных упражнений (с уровнем кислородного запроса 50—85 % МПК) — возрастает до 1—1,5 г ■ л-1 . Концентрация молочной кислоты значительно возрастает в первые 2—10 мин работы, а затем либо остается на прежнем уровне, либо снижается. Таким образом, максимальная концентрация молочной кислоты в крови наблюдается до тех пор, пока не установилось устойчивое состояние, создающее условия для аэробного ее окисления.
При выполнении упражнений с уровнем кислородного запроса более 85 % МПК концентрация молочной кислоты в крови постоянно увеличивается до максимальных значений. Концентрация молочной кислоты, которая не причиняет вреда организму хорошо тренированного человека, составляет 2—2,5 г ■ л-1 в крови. Дальнейшее увеличение концентрации молочной кислоты оказывает неблагоприятное воздействие на организм и тормозит процесс гликолиза.
Молочная кислота — сильная кислота, образующая при диссоциации значительное количество водородных ионов. Часть их может быть связана буферными системами клеток и крови, при этом в крови главную роль играет бикарбонатный, а в клетках — белковый буфер. Когда емкость буферных систем исчерпывается, происходит сдвиг активной среды в кислую сторону. В закислении среды участвуют и такие кислоты, как угольная фосфорная, пировиноградная и др. Однако роль молочной кислоты в этог.' процессе наиболее значительна. Между концентрацией молочной кислоть и величиной рН крови существует выраженная обратно пропорциональная зависимость. Как видно из рис. 147, максимальное значение концентрации молочной кислоты в крови в условиях напряженной мышечной деятельности достигает 20—25 ммоль ■ л'1 и более, а значение рН снижается от 7,4 в состоянии покоя до 6,9—6,8.
Снижение величины рН более чем на 0,2 по сравнению с уровнем покоя вызывает уменьшение активности многих ферментов, и в первую очередь фосфофруктокиназы, контролирующей ключевую реакцию глико-
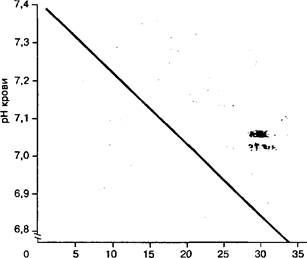
Рис. 147
Взаимосвязь между
изменениями
значения рН
и концентрации
лактата в крови при
напряженной
мышечной работе
лиза, поэтому общая скорость гликолиза снижается. Закисление среды
организма приводит также к нарушению деятельности нервных клеток и
развитию в них охранительного торможения, ухудшению передачи возбуж-
дения с нерва на мышцу, снижению АТФ-азной активности миозина и па-
дению скорости расщепления АТФ. Высокая концентрация молочной кис-
лоты в мышечных волокнах вызывает повышение в них осмотического дав-
ления, ведущего к набуханию их, сдавливанию нервных окончаний, в ре-
зультате чего могут возникать боли в мышцах. Многие спортсмены могут
вынести снижение рН крови до 6,8 и даже 6,5 (при изнеможении), однако
при этом наблюдаются тошнота, головокружение и сильные боли в мыш-
цах. Сдвиг величины рН крови в щелочную сторону возможен до 7,6, что
организм переносит без резких нарушений обменных процессов.
Избыток молочной кислоты в крови связывается бикарбонатным буфе-
ром, в частности его щелочным компонентом (ЫаНС03
):
+ НСОз + СНОНСОО" + Н+ -------- + СНзСНОНСОО" + НгСОз
XX
со2 + н2 о
В результате такого взаимодействия образуется так называемый из-
быток неметаболической углекислоты, которая не связана с процессами
биологического окисления. Она быстро распадается на С02
и Н2
0. Опре-
деляя долю неметаболического С02
в выдыхаемом воздухе, можно доста-
точно точно оценить степень усиления гликолитического процесса в рабо-
тающих мышцах.
Существует определенное соотношение между количеством выделен-
ного углекислого газа (УС02
) и потребляемого кислорода (1/02
), что назы-
вают дыхательным коэффициентом (ДК = 1/С02
/ У02
), который зависит от природы окисляемого энергетического субстрата. При окислении углеводов дыхательный коэффициент равен 1,0 (6С02
/ 602
= 1,0), при окислении жиров — 0,70, при окислении белков — 0,80, а при сбалансированной бел- ково-углеводно-жировой диете — около 0,75. Таким образом, по величине дыхательного коэффициента можно судить о характере окисляемых веществ и протекании окислительного процесса. Однако при напряженной мышечной работе дыхательный коэффициент может быть выше 1, что связано с появлением избытка молочной кислоты, увеличивающей образование и выделение С02
.
Мышечная работа вызывает изменение содержания в крови белков и продуктов их распада. Отмечается увеличение содержания белков в плазме крови (в частности, белков-ферментов) за счет их выхода из работающих мышц, а также изменяется соотношение между различными белками крови, увеличивается количество продуктов белкового распада — аминокислот, поступающих из мышечных клеток и печени, аммиака, мочевины. Изменения белкового обмена зависят от длительности работы. Так, при кратковременной работе выход белков из тканей в кровь незначителен, а при длительной работе, когда проницаемость клеточных мембран сильно изменяется, белок может проникать через клеточные мембраны почек и появляться в моче. Уровень аммиака особенно возрастает в случае, когда не устанавливается устойчивое состояние метаболизма, а также при длительной утомительной мышечной нагрузке. Длительная работа приводит также к увеличению содержания в крови мочевины.
4) Показать значение экспресс-методов биохимическом контроле в оценке функционального состояния спортсмена
Немаловажное значение в биохимическом обследовании имеют используемые методы определения показателей метаболизма, их точность и достоверность.
В настоящее время в практике спорта широко применяются лабораторные экспресс-методы определения многих (около 60) различных биохимических
показателей в плазме крови с использованием портативного прибора
швейцарской фирмы «Доктор Ланге» или других фирм. К экспресс-методам определения функционального состояния спортсменов относится также предложенный академиком В.Г. Шахба-зовым новый метод определения
энергетического состояния человека, в основу которого положены изменения
биоэлектрических свойств ядер эпителиальных клеток в зависимости от
физиологического состояния организма. Данный метод позволяет выявить нарушение гомеостаза организма, состояние утомления и другие изменения при мышечной деятельности.
Контроль за функциональным состоянием организма в условиях учебно-
тренировочного сбора можно осуществлять с помощью специальных
диагностических экспресс-наборов для биохимического анализа мочи и крови.
Основаны они на способности определенного вещества (глюкозы, белка,
витамина С, кетоновых тел, мочевины, гемоглобина, нитратов и др.)
реагировать с нанесенными на индикаторную полоску реактивами и изменять
окраску. Обычно наносится капля исследуемой мочи на индикаторную полоску
«Глюкотеста», «Пентафана», «Меди-теста» или других диагностических тестов
и через 1 мин ее окраска сравнивается с индикаторной шкалой, прилагаемой
к набору.
Одни и те же биохимические методы и показатели могут быть использованы
для решения различных задач. Так, например, определение содержания
лактата в крови используется при оценке уровня тренированности,
направленности и эффективности применяемого упражнения, а также при
отборе лиц для занятий отдельными видами спорта.
В зависимости от решаемых задач изменяются условия проведения
биохимических исследований. Поскольку многие биохимические показатели у
тренированного и не тренированного организма в состоянии относительного
покоя существенно не различаются, для выявления их особенностей проводят
обследование в состоянии покоя утром натощак (физиологическая норма), в
динамике физической нагрузки либо сразу после нее, а также в разные
При обследовании спортсменов применяются различные типы тестирующих
физических нагрузок, которые могут быть стандартными и максимальными
(предельными).
5. Дать биохимическую характеристику заллинга
Заллинг – это подвид спортивного туризма (дист. - пешеходные), проводящийся исключительно в закрытых помещениях, подписанный отдельным документом Росспорта.
Эти дистанции находятся примерно в таком промежуточном интервале 1,5-4 мин. Анаэробный процесс.
Гликоген
Глюкоза Креатин 
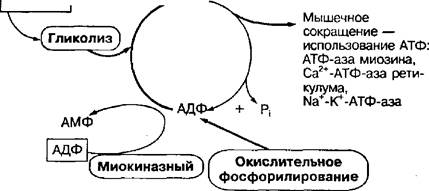
К анаэробным механизмам относятся:
• креатинфосфокиназный (фосфогенный или алактатный) механизм, обеспечивающий ресинтез АТФ за счет перефосфорилирования между креатинфосфатом и АДФ;
• гликолитический (лактатный) механизм, обеспечивающий ресинтез АТФ в процессе ферментативного анаэробного расщепления гликогена мышц или глюкозы крови, заканчивающегося образованием молочной кислоты, поэтому и называется лактатным;
• миокиназный механизм, осуществляющий ресинтез АТФ за счет реакции перефосфорилирования между двумя молекулами АДФ с участием фермента миокиназы (аденилаткиназы).
Анаэробные механизмы являются основными в энергообеспечении кратковременных упражнений высокой интенсивности, а аэробные — при длительной работе умеренной интенсивности.
Преобладают гликолитический механизм ресинтеза АТФ.
Энергетическое обеспечение работы в зоне субмаксимальной мощности осуществляется в основном за счет анаэробного гликолиза, что приводит к большому накоплению молочной кислоты в крови (концентрация ее может достигать 2,5 г ■ л1
и более). Кислородный запрос при такой работе может достигать 20—40 л, а уровень энергетических затрат может в 4—5 раз превышать максимум аэробного механизма энергообразования. К концу работы возрастает доля аэробных реакций в ее энергообеспечении. Кислородный долг в этой зоне мощности наиболее значителен по абсолютным значениям (до 20 л) и составляет 50—90 % кислородного запроса. Усиливается мобилизация гликогена печени, уровень глюкозы в крови может
8-09-2015, 19:52