1) нейроны, которые возбуждаются светом, падающим на центр рецептивного поля, но затормаживаются, если свет падает на его периферию;
2) нейроны, которые затормаживаются светом в центре рецептивного поля и возбуждаются при действии света на его края.
Одновременная реакция нейронов первого и второго типов лежит в основе появления одновременного контраста за счет подчеркивания края изображения их антагонистическими рецептивными полями.
Размеры рецептивных полей, равно как и их функциональные свойства, зависят от расположения фоторецепторов данного поля на сетчатке относительно центральной ямки (эксцентриситет ). Величина рецептивных полей растет от области центральной ямки к периферии сетчатки. Это является следствием структурной организации связей элементов сетчатки. Так, в середине сетчатки, в области центральной ямки, где плотность колбочек максимальная, одна колбочка через отдельную биполярную клетку соединяется с отдельной ганглиозной клеткой. Таким образом, рецептивные поля ганглиозных клеток, связанные с центром сетчатки очень узкие и не перекрываются. На периферии, где, в основном, находятся палочки, отмечаются широкие рецептивные поля: множество рецепторов связано с одной ганглиозной клеткой.
Функционально острота зрения зависит и от свойств рецептивных полей: острота зрения — возможность различения двух соседних точек — максимальна для узких рецептивных полей центральной ямки. В то же время слабые сигналы с периферии сетчатки выделяются зрением, благодаря взаимодействию широких перекрывающихся рецептивных полей за счет пространственной суммации раздражителей.
При освещении сетчатка генерирует электрические потенциалы, которые называют электроретинограммой . Эта суммарная электрическая реакция отражает процессы возбуждения различных нервных структур: волна возникает во внутренних сегментах фоторецепторов; волна является результатом возбуждения биполярных и амакриновых клеток сетчатки; волна связана с пигментным эпителием; волна, является реакций горизонтальных клеток сетчатки на выключение света.
Таким образом, волны электроретинограммы имеют своим источником все клеточные элементы сетчатки, кроме ганглиозных.
В сетчатке обнаружены клетки, для которых наиболее эффективны цвета-антагонисты красный и зеленый, а также клетки, для которых пару антагонистов составляют желтый и синий или зеленый и синий. Объяснение антагонистического действия цветов на ганглиозные клетки состоит в том, что из трех типов колбочек два всегда связаны с одним нейроном, а часть колбочек имеет возбудительные синапсы, другая часть — тормозные.
Сетчатку, по сложности организации, часто рассматривают как часть мозга, расположенную на периферии. Здесь с фоторецепторами связаны несколько слоев нейронов, формирующих афферентный поток, который идет к подкорковым и корковым центрам зрительной системы. Горизонтальные и биполярные клетки сетчатки не генерируют потенциалов действия, основной формой их активности являются градуальные гиперполяризации и деполяризации. Ганглиозные клетки генерируют потенциалы действия, которые проводятся по их длинным аксонам, составляющим зрительный нерв.
Зрительный нерв содержит около 800 тысяч волокон ганглиозных клеток сетчатки. Зрительные нервы обоих глаз перекрещиваются в области основания черепа, где около полумиллиона волокон зрительного нерва переходят на противоположную сторону. Остальные 300 тысяч волокон вместе с перекрещенными аксонами второго зрительного нерва образуют зрительный тракт.
Нервные волокна зрительного тракта подходят к четырем структурам мозга: ядрам верхних бугров четверохолмия — средний мозг; ядрам латерального коленчатого тела — таламус; супрахиазмальным ядрам гипоталамуса; к глазодвигательным нервам.
Ядра верхних бугров четверохолмия и латерального коленчатого тела являются конечными пунктами двух параллельных путей от ганглиозных клеток сетчатки: одна ветвь аксона ганглиозной клетки идет в латеральное коленчатое тело, другая — в верхнее двухолмие. Обе ветви сохраняют упорядоченную проекцию сетчатки. От переднего двухолмия после переключения сигналы идут к крупному ядру таламуса — подушке.
Аксоны клеток латерального коленчатого тела, проходящие в составе зрительной радиации, проецируются к клеткам первичной зрительной коры (поле 17 или стриарная кора). Проекция зрительной ямки сетчатки — зоны максимальной остроты зрения — в 35 раз больше проекции участка такого же размера на периферии сетчатки. Клетки поля 17 (стриарной коры) связаны с полями 18 и 19 (престриарная кора), так называемыми вторичными зрительными зонами. От этих зон идут проекции к подушке таламуса, куда поступает информация от верхних бугров четверохолмия. Кроме того, зрительные пути прослеживаются к лобной коре, они примыкают к ассоциативной коре.
Клетки латерального коленчатого тела, получающие основную афферентацию от сетчатки, имеют простые концентрические рецептивные поля, как и ганглиозные клетки. Здесь проявляется бинокулярное взаимодействие: волокна от обоих глаз распределены топографически правильно, послойно. В то же время небольшая часть клеток латерального коленчатого тела активируется от обоих зрительных нервов.
Нейроны зрительной коры уже имеют не концентрические, а почти прямоугольные зрительные поля, некоторые из нейронов реагируют на определенную ориентацию (наклон) полосы — светлой или темной (рис. 1.4).
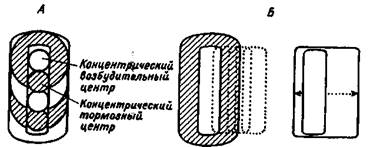
Рис. 1.4. Концентрические рецептивные поля в сетчатке и подкорковых зрительных центрах (А), прямоугольные и сложные рецептивные поля в зрительной коре (Б).
В зрительной коре существуют два функционально различных типа клеток: простые и сложные. Простые клетки имеют рецептивное поле, состоящее из возбудительной и тормозной зоны, которые можно предсказать на основе исследования реакции клетки на маленькое световое пятно. Структуру рецептивного поля сложной клетки невозможно установить сканированием светового пятнышка. Они служат "детекторами" угла, наклона или движения линий в поле зрения. В коре уже совершенно отчетлива бинокулярная конвергенция: в одной точке представлены симметричные поля зрения — справа и слева.
Близко расположенные клетки зрительной коры "видят" только небольшую часть поля зрения. Лежащие друг под другом нейроны одной колонки коры реагируют на один и тот же стимул, оптимальный по ориентации, наклону и направлению движения. В одной колонке могут располагаться как простые, так и сложные клетки. Простые клетки найдены в III и IV слоях, где заканчиваются таламические волокна. Сложные клетки расположены в более поверхностных слоях коры 17 поля. В полях 18 и 19 зрительной коры простые клетки являются исключением, здесь расположены сложные и сверхсложные клетки. Последние реагируют, например, только на стимулы определенной ширины, длины и ориентации.
Итак, от уровня к уровню зрительной системы происходит усложнение рецептивных полей нейронов. Все рецептивные поля организованы в виде возбудительных и тормозных зон. Концентрические рецептивные поля, характерные для сетчатки и латерального коленчатого тела, уже не встречаются в коре. В зрительной системе, как и в других сенсорных системах, чем выше синаптический уровень, тем строже ограничены функции отдельных нейронов — детекторов свойств.
Для успешной работы системы распознавания зрительных образов очень важны движения глаз . Известно, что глаз человека приводится в движение шестью наружными мышцами. Относительно координат головы глаза двигаются горизонтально, вертикально и вокруг своей оси. Если оба глаза двигаются в одном направлении, такие движения называются содружественными . При переводе взгляда с ближней точки на дальнюю осуществляются дивергентные движения. При наклоне головы в сторону наблюдаются небольшие вращательные движения глаз.
При взгляде на любой предмет глаза двигаются от одной точки фиксации к другой быстрыми скачками — саккадами . Длительность саккад от 10 до 80 мс, длительность периодов фиксации 150-300 мс. Медленные движения глаз реализуются при слежении за движущимися объектами — следящие движения.
Движения глаз управляются центрами, которые находятся в области ретикулярной формации мозга и среднего мозга, в верхних буграх четверохолмия и в претектальной области. Все эти подкорковые центры координируются сигналами из зрительной, теменной и лобной коры, ответственными за программирование движений тела и оценки его положения в пространстве. Для наиболее тонкой регуляции глазодвигательных функций весьма существенны влияний мозжечка, сравнивающего тонический и фазный компоненты движения при ориентации и пространстве.
В процессе зрительного восприятия, особенно при слежении за движущимся объектом, возникает оптический нистагм , вызываемый движущимися оптическими стимулами и состоящий из чередовании саккад и медленные следящих движений. Движения глаз имеют огромное значение для восприятия: при неподвижном глазном яблоке восприятие изображения пропадает в связи с разложением пигмента и адаптацией фоторецепторов.
Координированные движения глаз обеспечивают объединение информации, идущей от обоих глаз в центры мозга. Особое значение для восприятия и координации движений играют нейроны переднего двухолмия. Они организованы в колонки, которые воспринимают сигналы, поступающие от одних и тех же участков полей зрения: активность нейронов мою отдела мозга, на которых конвергирует импульсация от правого и левого глаза, является пусковым механизмом для глазодвигательных нейронов. На высших уровнях зрительной системы параллельно функционируют две системы анализа: одна определяет место предмета и пространстве, другая описывает его признаки. Конечные результаты параллельных процессов интегрируются и возникает законченный зрительный образ внешнего предметного мира.
2 СОМАТОВИСЦЕРАЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
2.1 Психофизика кожной механорецепции
Пороги субъективная интенсивность тактильных стимулов
Абсолютный порог для тактильного стимула можно измерить калиброванными волосками по методу, который ввел в психофизику в XIX в. немецкий физиолог Макс фон Фрей (Max von Frey). В современных стимуляторах высокоточные движения раздражающего стерженька обеспечиваются электромагнитом. Стерженек обеспечивает воздействие градуированной интенсивности с заданной временной характеристикой, например одиночные импульсы или ритмичные нажимы с синусоидально изменяющейся интенсивностью (вибрации).
Пороги восприятия, определенные волосками Фрея и вибрациями (200 Гц), генерируемыми электромагнитным стимулятором, приведены для разных частей тела в табл. 2.1.
Таблица 2.1. Пороги восприятия механической стимуляции кожи человека
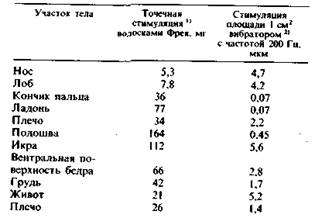
Кончики пальцев воспринимают вибрацию амплитудой менее 1 мкм. Относительная чувствительность разных частей тела весьма неодинакова. Кроме того, разные по своей природе стимулы демонстрируют неодинаковую чувствительность одного и того же места (табл. 2.1). Это объясняется тем, что околопороговая вибрация и волоски Фрея стимулируют разные типы механорецепторов.
Пространственное разрешение тактильных стимулов
Для получения меры пространственного paзрешения пригодны различные тесты по распознаванию пространственных деталей тактильных стимулов. В одном из часто применяемых тестов используя циркуль, двумя кончиками которого одновременно прикасаются к коже (рис. 2.1) для определения двухточечного порога.
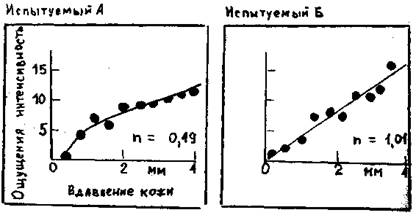
Рис. 2.1. Психофизические опыты для оценки интенсивности стимула. Испытуемый выражает ощущаемую им силу давления на ладонь (здесь – длительностью 1с.) числом на произвольно выбранной шкале (ось ординат). Объективную интенсивность стимула (ось обсцисс) меняли в случайном порядке. Кривые – степенные функции, соответствующие полученным данным с показателем степени n, отмеченным графиках.
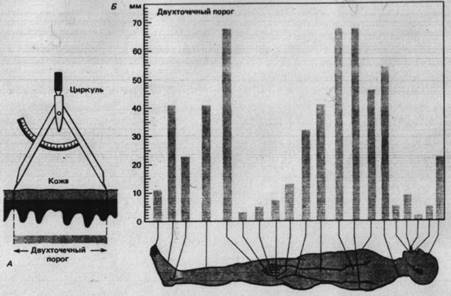
Рис. 2.2. Двухточечный порог кожи. А. Кончики циркуля, разведенные на разные расстояния, неоднократно прикасаются к коже. Двухточечный порог - наименьшее расстояние, при котором испытуемый может определить, чтo стимулируются две точки, а не одна. Б. Изменчивость двухточечного порога кожи в зависимости oт участка тела человека.
Испытуемый должен, не глядя, ответить, в одной или в двух точках до него дотронулись. Двухточечный порог на разных частях тела неодинаков (рис. 2.2 Б). Это обусловлено особенности иннервации разных областей кожи и связанным с ней характером соединения афферентных волокон в ЦНС.
2.2 Кожные механорецепторы
Исследования на человеке и животных привели к хорошему пониманию физиологической функции гистологического строения механорецепторов кожи и обнаружили между ними корреляцию. Кожа млекопитающих содержит несколько основных типов механорецепторов.
Типы низкопороговых механорецепторов кожи
Нейрофизиологические опыты показали, что лишенная волос кожа крыс, кошек, обезьян и людей одержит три основных типа механорецепторов с афферентами группы II (А![]() ). Их называют рецепторами МА, БА и ТП. МА означает медленно адаптирующийся: при долговременном воздействии механического стимула (давления; например, давления веса тела на подошву) такой рецептор посылает по своему афферентному волокну непрерывный поток импульсов. БА – это быстро адаптирующийся рецептор, реагирующий только на механические стимулы, которые изменяются во времени. ТП означает тельца Пачини
, гистологически выявленные структуры, функционирующие как очень быстро адаптирующиеся механорецепторы.
). Их называют рецепторами МА, БА и ТП. МА означает медленно адаптирующийся: при долговременном воздействии механического стимула (давления; например, давления веса тела на подошву) такой рецептор посылает по своему афферентному волокну непрерывный поток импульсов. БА – это быстро адаптирующийся рецептор, реагирующий только на механические стимулы, которые изменяются во времени. ТП означает тельца Пачини
, гистологически выявленные структуры, функционирующие как очень быстро адаптирующиеся механорецепторы.
Рецептивные поля и плотность иннервации механорецепторов
Область, в которой стимул данной интенсивности способен возбудить механочувствительное афферентное волокно, называется его рецептивным полем . Оно приблизительно соответствует анатомической протяженности всех окончаний этого волокна. Например, один афферент связан с двумя-тремя тактильными тельцами оволосненной кожи, а все 30-50 дисков Меркеля такого тельца иннервируются одной коллатералью. Концевые разветвления гораздо протяженнее у рецепторов волосяных фолликулов: каждый афферент может отходить от многих фолликулов, и каждый фолликул иннервируется несколькими афферентными волокнами.
Какие свойства рецепторов определяют пространственное разрешение, измеряемое двухточечным порогом? Согласно результатам различных исследований, величина рецептивного поля здесь не главное; решающую роль играет плотность иннервации (т. е. число афферентных волокон на 1 см2 поверхности кожи). Валлбо с сотрудниками, проведя микронейрографические измерения у людей, пришли к выводу, что пространственному разрешению на разных участках кисти соответствует только плотность афферентов БА и MA. Поскольку человек, как известно, лучше всего распознает предметы на ощупь подвижными пальцами, можно предположить главную роль в этом рецепторов БА.
2.3 Психофизика терморецепции
Терморецепция (температурная чувствительность) кожи включает два качественных типа – чувство холода и чувство тепла. Известно, что в коже человека есть специфические холодовые и тепловые точки, в каждой из которых можно вызвать ощущение только холода или только тепла. Холодовых точек больше, чем тепловых.
Например, на 1 см2 ладони 1-5 холодовых и только 0,4 тепловых. Наибольшая плотность этих точек обнаружена в самой чувствительной к температуре области – на лице; здесь на 1 см2 кожи 16-19 холодовых точек, а одиночных сенсорных точек, реагирующих на тепло, не обнаруживается. Восприятие температурных стимулов, включая сопровождающие их регуляторные реакции, обладает аффективным действием. Иными словами, они могут казаться приятными или неприятными: человек может мерзнуть или чувствовать освежающую прохладу, согреваться или мучиться от жары.
Психофизическое исследование терморецепции
Существует температурный диапазон, в котором при постоянстве температурного стимула мы не ощущаем ни тепла, ни холода; иначе говоря, в этой игральной зоне температурная чувствительность полностью адаптирована. За пределами нейтральной зоны устойчивые температурные ощущения возникают даже при постоянной температуре (ноги могут мерзнуть часами). Верхний и нижний пределы нейтральной зоны для участка кожи площадью 15 см2 равны соответственно 36 и 30 °С.
Устойчивые ощущения тепла и холода
Продолжительное ощущение тепла при температуре кожи выше 36 °С тем сильнее, чем выше эта температура. При температуре около 45 °С чувство тепла сменяется болью от горячего. Когда обширные области охлаждаются до температуры ниже 30 °С, возникает устойчивое ощущение холода; боль от холода возникает при температуре кожи 17 °С и ниже.
Динамические температурные ощущения
Температурные ощущения, испытываемые при изменении температуры кожи, в основном определяются тремя параметрами: ее исходной температурой, скоростью изменения последней и размерами участка, на который действует стимул.
Прохладную кожу (скажем, при 28 °С) нужно только слегка охладить (меньше чем на 0,2 °С), чтобы превратить устойчивое чувство холода в ощущение «стало холоднее». Но тот же участок кожи надо нагреть почти на 1 °С для появления чувства тепла. Аналогичным образом, при исходной температуре кожи 38 °С уже незначительное нагревание (менее чем на 0,2 С) вызывает ощущение «стало теплее», между тем как для появления чувства холода надо охладить кожу приблизительно на 0,8 °С.
Если скорость изменения температуры превышает примерно 5 °С/мин, это мало влияет на тепловой или холодовой пороги; при более медленных ее изменениях оба будут постепенно повышаться. Например, если кожа охлаждается на 0,4°С/мин, начиная от температуры 33,5 °С, для появления чувства холода понадобится 11 мин; за это время температура упадет на 4,4 °С. Когда охлаждение идет очень медленно, человек может не заметить, как обширные участки кожи стали совсем холодными (при одновременной потере тепла телом), особенно если его внимание отвлечено чем-то другим. Предположительно этот фактор действует, когда человек простужается.
2.4 Терморецепторы
Реакции при постоянной температуре кожи
При переходе примерно от 30 до 43 °С частота импульсации у тепловых рецепторов возрастает, а затем резко падает; в пороговой области ощущения боли от горячего (приблизительно 45 °С) тепловые рецепторы обычно невозбудимы. Частота импульсации холодовых рецепторов повышается по мере падения температуры от 33-40°С и достигает максимума при 17-26 °С (у разных рецепторов). Из-за этого максимума каждая частота разрядов связана с двумя температурами, но тем не менее ЦНС может правильно распознавать холод и на отрезке ниже 17°С.
Реакции при изменении температуры кожи
Сравнение показывает, что частота импульсации зависит не только от температуры (пропорциональный рецептор), но и от скорости ее изменения (дифференциальный рецептор). Поведение тепловых рецепторов при изменениях температуры как в зеркале отражает поведение холодовых: при нагревании их импульсация растет выше стационарного уровня, а при охлаждении падает ниже него, после чего возвращается к этому уровню. Температурные пределы динамической чувственности терморецепторов отличаются от установленных для статической. Например, при охлаждении кожи ниже максимума статической импульсации холодового рецептора динамическая частота его импульсации повышается, несмотря на решение ее статического уровня.
Следовательно, в целом выгоднее иметь и тепловые, и холодовые терморецепторы, а не дородную их популяцию, поскольку это дает ДНС значительно более детальную информацию о термических процессах в коже.
2.5 Висцеральная чувствительность
Спинномозговые и некоторые черепномозговые нервы содержат афференты, идущие от внутренних органов. Главные периферические рвы, в которых они проходят, - блуждающий, внутренностньй и тазовый. Эфферентные волокна их нервов относятся к парасимпатической и симпатической системам, а афферентные – к другой.
Информация, проводимая висцеральными афферентами, используется главным образом для вегетативных регуляторних процессов, управляющих кровообращением, дыханием, пищеварением, электролитическим и водным балансами.
Активность висцеральных афферентов до которой степени воспринимается сознательно. Это могут быть ощущения, обусловленные непосредственно
8-09-2015, 22:06