Сердечно-сосудистая система
В регуляции давления и объема крови участвуют механорецепторы в аорте, сонной артерии и предсердиях. Их непрерывную активность мы не осознаем. Однако в некоторых особых ситуациях нами ощущается собственная сердечная деятельность, например при сильном физическом или психическом напряжении. В этом восприятии сердцебиений, вероятно, также участвуют механорецепторы в коже и мышцах, возбуждаемые значительными изменениями формы и положения сердца, а также пульсацией артерий. Речь идет о непрямом восприятии активности висцеральных афферентов через запускаемые ими рефлексы.
Легочная система
Афференты, регулирующие дыхание, - это прежде всего хеморецепторы в артериальной системе и, возможно, также в мозге, возбуждающиеся понижением парциального давления О2 и повышением его у СО2 . Активность этих рецепторов тоже, как правило, не ощущается. Только когда парциальное давление СО2 резко возрастает и/или воздухоносные пути перекрываются (например, при астме), человек чувствует, как задыхается. Снижение парциального давления О2 из-за недостатка кислорода во вдыхаемом воздухе (например, при восхождения на высокую гору или при падении давления в кабине самолета) не ощущается, даже если оно настолько сильно, что вызывает полубессознательное состояние. Следовательно, непосредственное участие хеморецепторов, анализирующих кровоток, в появлении одышки маловероятно; воспринимается скорее недостаток двигательного выполнения дыхательной программы. Ритмичные дыхательные движения осознаются, вероятно, благодаря возбуждению многих механорецепторов в грудном скелетно-мышечном аппарате и коже.
Желудочно-кишечная система
По своему эмбриологическому происхождению желудочно-кишечный тракт-часть поверхности тела. Их функциональное соответствие заключается в том, что при переработке проглоченной пищи на желудок и кишки действует множество стимулов, возникающих во внешней среде. Как и следовало ожидать, механические, тепловые и химические стимулы воспринимаются желудочно-кишечной системой сильнее, чем другими внутренними органами. Особенно отчетливо она реагирует на растяжение; это можно доказать экспериментальным раздуванием баллона, введенного в пищеварительный тракт при помощи катетера. Растяжение желудка вызывает чувство наполнения, или сытости, растяжение кишечника – чувство закупорки (как при кишечных газах). В таких опытах с надуваемым баллоном стимулы иногда ошибочно локализуются испытуемым в зонах Геда на поверхности кожи. Сильная стимуляция желудочно-кишечной системы растяжением ощущается как боль, особенно когда вызывает контрактуру гладкой мускулатуры.
Тепловые и химические стимулы воспринимаются главным образом в пищеводе и прямой кишке. Судя по наблюдениям, сделанным в то время, когда операции на органах брюшной полости производились без наркоза, прикосновение к желудочно-кишечным структурам и их пальпация не осознаются.
Почечная система
В области почек и мочеточников человек способен ощущать только боль. Однако, если мочевой пузырь настолько наполнен, что растягивается, возникает потребность его опорожнить, обусловливающая произвольное мочеиспускание.
2.6 Проприоцепция
Восприятие позы и движения нашего собственного тела называется проприоцепцией (а также глубокой, или кинестетической, чувствительностью). Поза определяется углом расположения костей в каждом суставе, устанавливаемым либо пассивно (внешними силами), либо активно (мышечным сокращением). Рецепторы проприоцепции находятся не только в суставах, но также в мышцах сухожилиях и коже. С их работой сочетаются сигналы от вестибулярного органа, что позволяет определять положение тела в поле земного притяжения. Все эти проприоцепторы участвуют также и в нашей сознательной и бессознательной двигательной активности.
Типы проприоцепции
Чувство позы. Даже с закрытыми глазами мы осознаем углы между костями в наших суставе а значит, и относительное положение частей тела. Эту способность легко продемонстрировать на простых опытах, выполняемых испытуемыми с закрытыми глазами. Если конечности на одной стороне тела пассивно или активно придано какое-нибудь положение (например, рука согнута в локте), испытуемый может точно воспроизвести его конечностью на другой стороне. Если попросить его показать на себе определенный участок тела, он сможет это сделать с очень небольшой ошибкой.
Чувство движения. При изменении положения сустава мы воспринимаем и направление, и скорость этого движения. Амплитудный порог такого восприятия зависит от угловой скорости.
Чувство силы. Благодаря ему мы воспринимаем степень мышечного усилия, нужного для выполнения движения или поддержания позы (например, для разных гравитационных нагрузках), и достаточно хорошо оцениваем вес предметов, поднимая их рукой. Сравнивая вес двух предметов, одновременно поднимаемых обеими руками, мы воспринимаем разницу уже на уровне 3-10%. Если сравниваемые предметы положить на руки, а те на подставку, оценки веса, основанные теперь на афферентной информации от кожных рецепторов, будут значительно менее точны.
Проприоцепторы
Сознательное восприятие движения и положения суставов, по-видимому, обусловлено одновременным действием различных механорецепторов: суставных и кожных рецепторов, мышечных веретен, сухожильных органов. С этим согласуется и то, что соматосенсорное ядро таламуса (вентробазальный юмплекс) и соматосенсорная кора получают информацию от всех перечислении выше их типов. В последние годы возникли некоторые разногласия по поводу относительного участия в восприятии каждого из них.
Долго считалось, что проприоцепция обусловлена главным образом суставными рецепторами. Эти механорецепторы в суставных капсулах кодируют различные положения или движения суставов. Однако их предполагаемая особая роль в проприоцепции стала оспариваться, когда было установлено, что больные с искусственными суставами различает их положение почти так же, как обычные люди. Более поздние психофизические опыты по восприятию положения и движения суставов подтвердили гипотезу, согласно которой важнейшую роль в нем играют мышечные веретена. Воздействуя на мышцы и сухожилия вибрацией, активирующей главным образом мышечные веретена и сухожильные органы, можно вызывать иллюзии движения в суставах, сбивающие испытуемых с толку.
Вывод, вытекающий из многочисленных психофизических исследований и клинических наблюдений, состоит в том, что для проприоцепции ЦНС использует всю доступную ей нервную информацию. По всей вероятности, афферентные сигналы от мышечных веретен, сухожильных органов, суставных и кожных рецепторов интегрируются, а это приводит к сложному восприятию положения и движения тела. Эфферентная двигательная информация, например команды от двигательной коры, скорее всего, также используется в этом процессе в виде копии эфферентации.
2.7 Функциональный и анатомический обзор центральной соматосенсорной системы
Центральную переработку сигналов от периферических рецепторов можно рассматривать на трех функциональных уровнях - афферентной, интегративной и эфферентной подсистем. Афферентный, или сенсорный, уровень соматовисцеральной системы связан со спинным мозгом, стволом мозга, таламусом и корой больших полушарий. Действуют различные проводящие пути, позволяющие перерабатывать информацию параллельно, используя ее для выполнения разных функций. Периферическая сенсорная поверхность (совокупность всех рецепторов вне ЦНС) проецируется на таламус и кору таким образом, что упорядочение (соматотопически) картируется в ЦНС. Следующие этапы центральной нервной переработки информации соответствуют интегративному и эфферентному уровням. Ассоциативная и лимбическая системы выполняют главным образом интегративные функции. Одна из важнейших задач этих систем—сопряжение сигналов от различных сенсорных систем с информацией, хранящейся в памяти. Двигательная и вегетативная системы выполняют эфферентные функции. Сложные взаимодействия трех функциональных подсистем, по-видимому, лежат в основе поведения. Оно в присутствии сенсорных стимулов делится на восприятия и реакции с пятью компонентами: когнитивным (включающим осознанное различение), аффективным (связанным с эмоциями), мотивационным (связанным с побуждениями, или драйвами), двигательным и вегетативным.
Дихотомия в восходящей центральной соматовисцеральной системе Афферентные нервные волокна от туловища и конечностей идут в спинной мозг по спинномозговым нервам, а от лица – в ствол мозга по тройничному нерву. Сигналы от рецепторов различным образом используются в обоих этих центральных отделах для двигательных и вегетативных рефлексов, а также передаются в высшие отделы головного мозга по системам восходящих волокон. На рисунке 2.3 для общей ориентации в таких связях дана сильно упрощенная их схема.
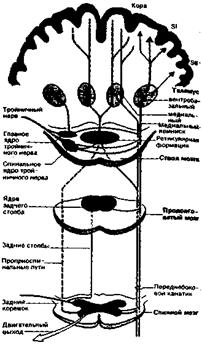
Рис. 2.3. Анатомическая организация главных функциональных элементов соматосенсорной системы (схема).
Соматосенсорная информация передается по двум главным системам восходящих трактов, различающихся как анатомически, так и функционально. Это системы:
- заднего столба, или лемнисковая;
- переднебокового канатика.
Главные связи в обеих системах служат для соединения каждой стороны тела с противоположной (контралатеральной) половиной головного мозга. Функционально система заднего столба связана с кожной механорецепцией и проприоцепцией, а система переднебокового канатика – главным образом с терморецепцией и ноцицепцией. Хотя в спинном мозгу обнаружены дополнительные восходящие пути соматосенсорной информации, полезно сохранять именно такое деление, основанное на двух главных системах трактов.
Последствия повреждения спинного мозга
Когда спинной мозг только «надорван» с одной стороны (например, при несчастном случае), прерывание трактов в его белом веществе вызывает характерный неврологический дефицит, синдром Броун-Секара. Он заключается в сенсорных нарушениях с обеих сторон тела ниже уровня повреждения с параличом произвольных движений на ипсилатеральной стороне. Сенсорная недостаточность с двух сторон тела неодинакова: испилатерально нарушено осязание (повышен, например, двухточечный порог, а контралатерально-восприятие боли и тепловых стимулов. В то же время ипсилатеральные ноцицепция и терморецепция, как и контралатеральное осязание, нормальные. Поэтому такой эффект называется диссоциированным сенсорным дефицитом.
Он обусловлен разрывом трактов в белом веществе. Блокада нисходящих двигательных путей (например, пирамидного тракта) вызывает ипсилатеральный паралич произвольных движений, повреждение заднего столба нарушает ипсилатеральное осязание, а переднебокового канатика – контралатеральные ноцицепцию и терморецепцию. Сенсорная информация от определенной области кожи идет по заднему столбу и переднему канатику с обоих сторон, но на уровне головного мозга в случае обоих этих сенсорных путей она воспринимается преимущественно контралатерально той стороне тела, на которой действует стимул.
Система заднего столба
Название этой системы отражает тот факт, что сенсорные сигналы передаются по заднему столбу (или канату), т.е. массивному пучку волокон в дорсальном белом веществе спинного мозга. Он состоит из прямых коллатералей толстых миелинизированных афферентов спинномозговых нервов. Другое название – лемнисковая система – обусловлено тем, что выше спинного мозга ее сигналы проводятся по медиальному лемниску (медиальной петле). Эта восходящая соматосенсорная подсистема соединяет спинномозговые и тройничные механорецептивные афференты с двумя анатомически и нейрофизиологически разделенными участками теменной коры – первой и второй соматосенсорными областями.
Афференты в системе заднего столба идут от низкопороговых кожных механорецепторов (МА, БА и ТП), мышечных веретен, сухожильных органов и суставных рецепторов. Проведение в этом восходящем пути быстрое, поскольку здесь только три синаптических переключения. Он состоит из следующих основных частей (рис. 2.3): задних столбов спинного мозга, ядер заднего столба в продолговатом мозгу (первый синапс), медиального лемниска, перекреста (переход путей на другую сторону), вентробазалъного ядра таламуса (второй синапс), корковых областей SI и SII (третий синапс). Тройничные коллатерали образуют синапсы в главном тройничном ядре ствола мозга, соответствующем ядрам заднего столба. После перекреста проводящий путь продолжается по медиальному лемниску к таламусу.
Особое свойство системы заднего столба – упомянутая выше соматотопическая организация, т.е. примерное сохранение топографических связей разных областей кожи (периферической сенсорной поверхности) во всех центральных зонах переключения. Это приводит к геометрически искаженному картированию, или проецированию, первых на вторые, подобные тому, которое свойственно зрительной и слуховой системам.
Система заднего столба особенно хорошо развита у приматов, т.е. у обезьян и человека. Она представляет собой анатомический субстрат тактильных ощущений (осязания) и проприоцепции, всех сознательных и бессознательных способностей, требующих различения пространственных и временных деталей механической стимуляции тела.
Сигналы от кожи, мышц и суставов, передаваемые по заднему столбу (особенно тактильные), дают важную информацию о выполнении движений. Поэтому у больных с повреждением заднего столба сильно понижена способность распознавать предметы, например, активно ощупывая их, или идентифицировать цифры, вычерчиваемые на коже. Двухточечный порог у них повышен.
Система переднебокового канатика
Эта система показана на рис. 2.3. Переднебоковой канатик включает в себя ряд трактов в вентральном белом веществе спинного мозга, содержащих восходящие волокна спинальных нейронов, тела которых находятся в сером веществе, главным образом в заднем роге. Тройничные аксоны, соответствующие переднебоковому канатику, идут от нейронов в спинальном тройничном ядре (рис. 2.3). Афферентные сигналы в переднебоковой канатик проводятся спинномозговыми и тройничными нервами от терморецепторов и ноцицепторов, а также от низкопороговых механорецепторов, в основном от расположенных в коже. Восходящие аксоны присоединяются к переднебоковому тракту, контралатеральному телам их клеток и входам их афферентных волокон в спинной мозг. Поскольку главных мест назначения два - ретикулярная формация с другими стволовыми областями мозга и таламус, различают спиноретикулярный и спиноталамический тракты. Передаваемая ими информация, по-видимому, в конце концов достигает многих отделов головного мозга, главным образом по полисинаптическим, медленно проводящим путям. Здесь нет ни четкой соматотопической организации, ни ясно очерченной проекции на кору, сравнимой с известной для системы заднего столба. Система переднебокового канатика рассматривается как анатомический субстрат терморецепции и ноцицепции и, кроме того, как часть неспецифичной системы.
Неспецифичная система
Приблизительно с 1950 г. в нейрофизиологии распространилось понятие неспецифичной системы как альтернативы четко определимым анатомически и физиологически специфичным системам различных сенсорных модальностей. В соматосенсорной модальности система заднего столба – специфичная. Неспецифичную систему рассматривали как субстрат ряда наблюдаемых в опытах на животных генерализованных peaкций типа:
- реакций возбуждения;
- изменения ЭЭГ;
- ритма сон/бодрствование;
- аффективных поведенческих реакций;
- изменений кровяного давления.
Они могут быть вызваны как сенсорными стимулами (особенно болевыми), так и электрической стимуляцией ретикулярной формации ствола мозга и медиальных таламических областей. Эти центральные области считались «сердцевиной» неспецифичной системы, получающей сенсорную информацию главным образом по переднебоковым трактам.
Исследования названного выше филогенетически древнего отдела головного мозга показывают, что здесь протекают многие жизненно важные интегративные и регуляторные процессы, отражающиеся на функциях спинного (например, симпатические реакции) и большого мозга (например, состояние тревоги). Однако достаточных свидетельств существования однородной неспецифичной системы нет. Напротив, с накоплением данных по физиологической, анатомической и биохимической дифферецировке этих отделов головного мозга сам такой термин становится все менее и менее приемлемым.
2.8 Передача соматовисцеральной информации в спинном мозгу
Дерматомы
Афференты от кожи, мышц, cycтавов и внутренних органов входят в спинной мозг задним корешкам в определенном пространственном порядке, т. е. согласно топологической организации. Кожные афференты каждого заднего корешка иннервируют ограниченную область кожи, называемую дерматомом. Смежные дерматомы сильно перекрываются из-за перераспределения пучков волокон при их распространении к периферии, особенно в плечевом и пояснично-крестцовом сплетениях. Один периферический нерв содержит волокна нескольких соседних задних корешков, а каждый задний корешок – волокна разных нервавов. В то время как перерезка периферического нерва вызывает сенсорный дефицит ограниченного участка тела, перерезка одного заднего корешка, хотя и обедняет иннервацию какой-то области, все же не вызывает в ней заметного сенсорного дефицита.
В скелетной мускулатуре эквивалентами дерматомов являются миотомы. Зона Геда внутреннего органа состоит из дерматомов, связанных со спинномозговыми сегментами, иннервирующими этот орган. Нейронная организация обеих совокупностей афферентов такова, что может вызывать отраженную боль.
Функциональные свойства спинальной соматовисцеральной системы
Соматовисцеральные афференты образуют синаптические связи с нейронами в заднем (дорсальном) роге спинного мозга. Некоторые толстые миелинизированные афференты (группы I ІI) также отделяют ветви (коллатерали), восходящие в составе спинного мозга. У заднего рога, где идет нейронная переработка информации, четыре выхода: длинные тракты, восходящие до головного мозга главным образом в переднебоковом канатике; восходящие и нисходящие проприоспинальные связи с соседними сегментами; сегментарные связи с мотонейронами и симпатическими нейронами (участие в спинальных двигательных и вегетативных рефлексах). Однако, по всей вероятности, один нейрон не образует все четыре вида связей сразу.
В восходящих трактах аксоны, идущие от одного сегмента, расположены бок о бок. Это создает соматотопическую слоистость; аксоны, присоединяющиеся к прочим во все более высоких сегментах, всегда оказываются со стороны серого вещества. Один из результатов такого расположения состоит в том, что при поверхностном повреждении переднебокового канатика (раной или опухолью) в шейной области первые сенсорные дефициты следует ожидать в нижней половине тела.
Нейроны переднебокового канатика
Хотя клинические данные указывают на участие переднебокового канатика в болевых и температурных ощущениях, он содержит всего лишь несколько аксонов специфичных ноцицептивных и терморецептивных нейронов. Большинство аксонов принадлежит здесь клеткам, возбуждаемым также слабыми механическими стимулами, т.е. мультирецептивным нейронам. Остается спорным вопрос, достаточно ли немногих специфичных ноцицептивных и терморецептивных нейронов для передачи соответствующих сигналов или дополнительная информация, требуемая головному мозгу для болевых и температурных ощущений, обеспечивается активностью мультирецептивных нейронов. Важное свойство заднего корешка как станции переключения – конвергенция афферентов от кожи и внутренних органов на одни и те же нейроны. Из-за нее афферентные сигналы от внутренностей (например, запускаемые желчными коликами) «ложно» локализуются больным на поверхности тела. Каждый внутренний орган связан с определенной областью кожи – своей зоной Геда. Другой элемент цепи заднего корешка – тормозные синапсы, которые служат для модификации афферентной информации. Это может принимать форму латерального торможения или регуляции сенсорного входа трактами, нисходящими от головного мозга. Активация таких тормозных механизмов применима для обезболивания.
Нейроны ядер заднего столба
Аксоны заднего столба оканчиваются ипсилатерально в его ядрах (клиновидном и тонком), находящихся в продолговатом мозгу. Здесь они образуют синапсы с крупными нейронами, посылающими аксоны в контралатеральный таламус по медиальному лемнисовому тракту и
8-09-2015, 22:06