- сохранение сенсорной специфики; на одном нейроне конвергируют афференты одинакового сенсорного типа;
- большой запас надежности синаптической передачи; даже одиночные импульсы aфферентного волокна вызывают постсинаптическую импульсацию;
- мелкие рецептивные поля;
- соматотопическая организация; пространственно упорядоченное представители кожи;
- афферентное торможение;
- нисходящие регулирующие входы, в частности, от коры головного мозга.
Эти свойства характерны и для последующих мест переключения в лемнисковой системе; они определяют качество и разрешающую способность ощущений, связанных с передаваемой здесь информацией.
2.9 Соматосенсорные функции ствола мозга
Ствол мозга, состоящий из продолговат мозга, моста и среднего мозга представляет собой место отхождения (или вхождения) большинства черепно-мозговых нервов. В нем содержится множество дискретных ядер, выполняющих преимущественно сенсорные, двигательные или регуляторные функции; между ними, по-видимому, проходят все восходящие и нисходящие пути связывающие головной мозг со спинным, а также большой мозг с мозжечком.
Тройничный нерв и его центральные связи
Голова иннервируется 12 парами черепно-мозговых нервов, большинство которых присоединяется к центральной нервной системе в стволе мозга и промежуточном мозгу. Пятый из этих нервов называемый тройничным из-за своих трех ветвей, включает афференты от лица и ротовой области (рис. 2.4).
Он иннервирует кожу, зубы, слизистую рта, язык и роговицу. Седьмой черепномозговой нерв (лицевой) тоже, но в меньшей степени, содержит головные соматосенсорные афференты. В языкоглоточном и блуждающем нервах есть висцеральные афференты от органов кровообращения, дыхания и пищеварения.
Сенсорные тройничные ядра и восходящие пути
Афференты тройничного нерва образуют синапсы в ядре спинального тракта и главном сенсорном ядре (рис. 2.4). Ядро спинального тракта функционально соответствует заднему рогу спинного мозга. Здесь механорецептивная, терморецептивная и ноцицептивная информация передается на аксоны, несущие ее к ретикулярной формации и таламусу, как и волокна переднебокового канатика, передающие туда информацию от спинного мозга. Все афференты, которые оканчиваются в главном сенсорном ядре, соответствующем ядрам заднего столба, идут от низкопороговых механорецепторов. Постсинаптические аксоны переходят на другую сторону и направляются к таламусу в составе медиального лемнискового тракта.
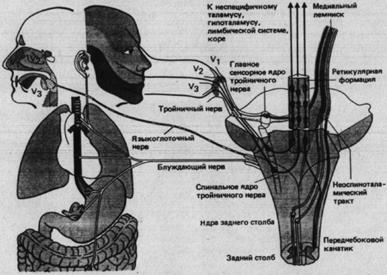
Рис. 2.4. Афференты и структуры соматовисцеральной системы в стволе головного мозга (справа - его схематичный вид сзади). К ним относятся афференты тройничного нерва (трех его ветвей V![]() , V
, V![]() и V
и V![]() ), блуждающего и языкоглоточного нервов, центральная афферентная система тройничного нерва и ее восходящие пути, ретикулярная формация и тракты, приходящие из спинного мозга, т.е. дорсальных столбов и переднебокового канатик. Центральное прохождение афферентов языкоглоточного и блуждающего нервов не показано.
), блуждающего и языкоглоточного нервов, центральная афферентная система тройничного нерва и ее восходящие пути, ретикулярная формация и тракты, приходящие из спинного мозга, т.е. дорсальных столбов и переднебокового канатик. Центральное прохождение афферентов языкоглоточного и блуждающего нервов не показано.
В стволе мозга информация, приносимая тройничными афферентами, интегрируется в двигательные рефлексы мускулатуры головы и во множество вегетативных рефлексов. Тройничная система осуществляет жизненно важные функции (особенно у млекопитающих) тактильного исследования окружающей среды, питания, издавания звуков и т.д. Она хорошо развита уже при рождении и проводит сенсорные сигналы, запускающие пищевое поведение, т.е. обеспечивает первое сенсорное знакомство ребенка с окружающей средой.
Ретикулярная формация
Ретикулярная формация – протяженная структура в стволе мозга (рис. 2.4)-важная интегративная область неспецифичной системы. Соматовисцеральные афференты попадают по спиноретикулярному тракту переднебокового канатика, а возможно, также по проприоспинальным (полисинаптическим) путям и соответствующим путям от ядра спинномозгового тройничного тракта. У ретикулярной формации есть афферентный вход и от других черепно-мозговых нервов. Кроме того, здесь много различных эфферентных связей: нисходящих к спинному мозгу и восходящих через неспецифичные таламические ядра к коре больших полушарий, гипоталамусу и лимбической системе.
Функции ретикулярной формации еще не вполне понятны. Ретикулярная формация участвует в:
- регуляции возбудимости коры: уровня осознания стимулов и реакций, ритма сон/ бодрствование (восходящая активирующая ретикулярная система);
- придании аффективно-эмоциональных аспектов сенсорным стимулам, особенно болевым, за счет передачи афферентной информации в лимбическую систему;
- двигательной регуляции, особенно связанной с так называемыми жизненно важными рефлексами (кровообращения, дыхания, глотания, кашля и чихания), требующими координации нескольких афферентных и эфферентных систем;
- регуляции позы и целенаправленных движений.
Этот перечень свидетельствует о невозможности четкого различения в ретикулярной формации цента сенсорной и интегративной систем. С другой стороны, недавно стало ясно, что некоторые ее области можно различать по анатомическим, функциональным и химическим свойствам (например, серотонинергических ядра шва и норадренергическое голубое пятно). Скорее всего, по мере расширения наших знаний о ретикулярной формации, представление о неспецифичной системе будет пересмотрено.
2.10 Таламус
Таламус считается входными воротами и распределительным пунктом, через которые все афферентные системы получают доступ к филогенетически более молодым церебральным структурам, обеспечивающим осознание сенсорных стимулов и сознательное целенаправленное поведение.
Анатомический и функциональный обзор
Правый таламус можно разделить на несколько функционально и/или анатомически различимых ядер, каждое из которых связано со своей корковой областью. Для обобщей ориентации сгруппируем эти ядра в следующие четыре функциональных класса:
- специфичные переключающие и перерабатывающие ядра кожных сенсорных органов, глаза и уха;
-ядра с преимущественно двигательными функциями;
-ядра с ассоциативными функциями;
- неспецифичные ядра без определенных корковых мишеней.
Таламические переключающие ядра сенсорных органов
Эти переключающие и перерабатывающие структуры связаны с корковой областью, отвечающей за их сенсорную модальность, и в свою очередь регулируются (возбуждаются и тормозятся) этой областью.
Неспецифичные ядра
Эта категория включает медиальные области, примыкающие к третьему желудочку мозга, а также интраламинарные ядра. Это высшие перерабатывающие и распределительные пункты для афферентной информации, конвертирующей на ретикулярной формации мозгового ствола. Сигналы от спинного мозга приходят сюда непосредственно по палеоспиноталамическому тракту и непрямо – по спиноретикулярному.
Двигательные ядра
Самое главное из них вентролатеральное (ВЛ), связывающее мозжечок и базальные ганглии с двигательной корой. Хирургические воздействия на соответствующие участки ВЛ могут ослабить двигательные расстройства (например, паркинсонизм).
Ассоциативные ядра
Эти части таламуса соединены с корой, но не могут быть отнесены к какой-либо определенной сенсорной системе; они принимают участие в интегратнвных функциях головного мозга.
Специфичное таламическое ядро соматосенсорной системы
Из-за своего анатомического положения этот участок лемнисковой системы называется вентробазальным ядром (ВБ) или вентробазалъным комплексом. В нем выделяют вентральное постеролатеральное (ВПЛ) и вентральное постеромедиальное (ВПМ) ядра. В ВПЛ находится нейронное представительство туловища и конечностей, а в ВПМ — лица. Главный проводящий путь, который ведет к ВПЛ, - это медиальный лемнисковый тракт, а к ВПМ тройнично-таламический тракт, начинающийся от главного сенсорного ядра тройничного нерва.
В опытах на наркотизированных животных установлены следующие функциональные особенности ВБ-нейронов:
-каждый из них обладает определенным рецептивным полем для механических стимулов кожи;
- рецептивные поля тем меньше, чем дистальнее они находятся на конечностях;
- соседние участки тела проецируются на соседние участки ВБ (пример соматотопической организации);
- каждый нейрон возбуждается главным образом рецептором одного типа (например, рецепторами МА или волосяных фолликулов в коже);
- импульсация нейрона усиливается с ростом интенсивности периферической стимуляции, т.е. интенсивность кодируется так же, как у кожных механорецепторов.
2.11 Соматосенсорные проекционные области в коре
Вентробазальный комплекс таламуса соединен как восходящими, так и нисходящими аксонами с двумя корковыми зонами – SI и SII (S означает «соматосенсорная область»). SI расположена на постцентральной извилине непосредственно позади глубокой центральной борозды, проходящей поперек полушария. SII лежит на верхней стенке боковой борозды, разделяющей теменную и височную доли. SI филогенетически моложе SII и очень важна у высших млекопитающих (особенно у приматов); она участвует во всех функциях соматосенсорной системы, зависящих от хорошего пространственно-временного различения стимулов. Некоторые из этих функций можно исследовать, например по методу двухточечного порога кожи (рис. 2.5)
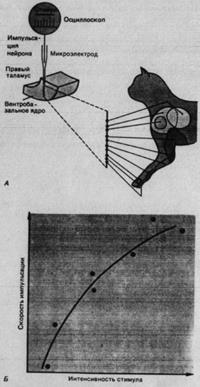
Рис. 2.5. Рецептивные поля нейронов в вентробазальном ядре таламуса. Микроэлектрод, проводимый через таламус наркотизированной кошки, прошел 10 нейронов, отвечающих на механическую стимуляцию кожи. Их рецептивные поля находились, как показано, на левой передней конечности.
Топографическая организация соматосенсорной коры
Соматотошпеское представительство периферии тела находится в ковтралатеральной SI, организовано поразительно подробно и было тщательно изучено. Вся поверхность тела здесь картирована причем у приматов даже имеются множествен проекции кисти. Сходное, хотя и менее четкое, картирование свойственно и SII, где оно частично билатерально. Соматотопическую opганизацию сенсорной коры изучают несколькими методами. У человека во время операции на головном мозгу применяют ее локальную электрическую стимуляцию с целью вызывать ощущения локализованные в соответствующих участках тела. Можно также с помощью радиоактивного ксенона измерять локальный корковый кровоток во время стимуляции кожи. У животных для картирования коркового представительства периферии тела использовали анализ вызванных потенциалов и запись активности одиночных нейронов.
Электрическая стимуляция коры мозга человека
При ряде нейрохирургических операций проводят локальную электрическую стимуляцию коры бодрствующего больного (для анестезии места разреза). Вызываемые этим ощущения воспринимаются так, как будто возникают на периферии. Систематическое обследование таким способом SI позволило получить картину, приведенную на рис. 2.6.
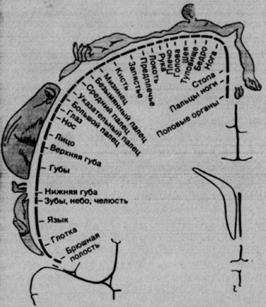
Рис. 2.6. Соматотопическая организация корковой зоны SI человека. Изображения над поперечным срезом мозга (на уровне постцентральной извилины) и их обозначения демонстрируют пространственное представительство поверхности тела в коре, установленное путем локальной электрической стимуляции мозга бодрствующих больных.
Этот «соматосенсорный гомункулус» – сильно искаженная карта периферической сенсорной поверхности с непропорционально крупными представительствами областей пальцев и pта. У человека они особенно густо иннервированы и, как показали психофизические опыты, характеризуются прекрасной пространственной разрешающей способностью, т. е. низкими двухточечными порогами. Очевидно, эти два факта причинно связаны, причем такая связь существует и в случае других сенсорных систем: чем больше рецепторов и центральных нейронов приходится на 1 мм2 сенсорной поверхности, тем лучше пространственное разрешение ею стимулов.
Переработка информации в нейронах соматосенсорной коры
Колонки корковых нейронов. При прохождении микроэлектрода перпендикулярно поверхности постцентральной извилины последовательно встречаемые им нейроны часто обладают идентичными или широко перекрывающимися рецептивными полями. Если же электрод перемещается под углом к этой поверхности, он проходит нейроны с соседними, но четко разграниченными рецептивными полями (в соответствии с соматотопической организацией). Из этих и других данных сделан вывод, что соматосенсорная кора, как и двигательная, организована в виде функциональных единиц-колонок нейронов, перпендикулярных ее поверхности.
Рецепторная специфичность колонок
Адекватная избирательная стимуляция, например, различных кожных рецепторов показывает, что нейроны одной колонки возбуждаются обычно рецепторами только одного типа. Очевидно, колонки – это функциональные единицы, соответствующие положению и сенсорной специфике периферических сенсорных нервных окончаний.
Простые и сложные нейроны
Характеристики импульсации простых корковых нейронов очень близки к таковым у связанных с ними рецепторов. Например, в SI есть нейроны, ведущие себя как рецепторы БА. Корковые нейроны, отвечающие на периферические стимулы импульсацией, явно отличающейся от той, что свойственна связанным с ними рецепторам, называются сложными. Этот термин охватывает разнообразные клетки. Среди них известны нейроны, которые реагируют только на стимулы, движущиеся линейно по поверхности кожи, и дают максимальный ответ на движение в определенном направлении. Они обнаружены в SI, SII и ассоциативных областях теменной коры.
Терморецептивные корковые нейроны, найденные у обезьяны, относятся к сложным. В отличие от периферических терморецепторов они реагируют либо только на изменения температур кожи, либо только на ее постоянную температуру. Кроме того, у них нет пика ответа при температуре около 26 о С, характерного для холодовых рецепторов. Другой тип сложных терморецепторных нейронов обнаружен в коре мозга крысы. Здесь частота импульсации «скачет» между максимумом и минимумом, когда температура соответствующих рецептивных полей кожи меняется приблизительно на 2 о С (пороговый детектор).
Эти примеры простых и сложных нейронов показывают, что корковая переработка сигналов приводит к нейронному представительству или отфильтровыванию информации о самых разнообразных параметрах периферических стимулов. Такая способность извлекать информацию, названная выделением признаков, особенно тщательно изучена в зрительной коре.
Эфферентные связи SI
От области SI, как и от всех прочих участков коры, отходит множество эфферентных аксонов. Как полагают они проводят в переработанном виде информации о периферических стимулах к другим частям центральной нервной системы. SI связана со следующими областями (указана и основная функция связи):
-двигательная кора: регуляция движения по обратной связи;
-теменные ассоциативные зоны: интеграция тельной и тактильной информации;
- контралатералъные SI и SII: интеграция билатеральной тактильной информации;
- таламус, ядра заднего столба, спинной мозг: эфферентная регуляция потока афферентной информации.
Корковая область SII значительно меньше SI. Она лежит у латерального конца постцентральной извилины, на верхней стенке сильвиевой борозды. Здесь соматотопическое представительство поверхности тела билатеральное. Рецептивные поля нейронных колонок обычно находятся на обеих сторонах тела, а их проекции часто расположены симметрично. Как полагают, SII играет особую роль в сенсорной и двигательной координации двух сторон тела (например, при хватании или ощупывании обеими руками).
Соматосенсорная кора и восприятие
Многие данные говорят о том, что SI необходима для точного тактильного различения и сознательного восприятия событий на поверхности и с высоким разрешением их силовых, пространственных и временных деталей. Впрочем, возбуждений SI необязательно осознается; например, периферическая стимуляция дает первичные вызванные потенциалы у наркотизированного или спящего человека сознание которого отключено.
Стимуляция коры человека
Локальная электрическая стимуляция SI или SII бодрствующего больного вызывает у него ощущения, которые часто идентичны возникающим при соответствующей стимуляции на периферии. Сенсорные ощущения возникают также (и только) при раздражении SII, зрительной и слуховой проекционных зон. Действие околопороговых точечных стимулов на SI описывалось больными, сходное с «естественной стимуляцией». При электрическом воздействии на кору возникали простые ощущения, специфичные для определенных рецепторов (вибрация, тепло, холод), ощущение перемещающихся по коже стимулов или движений суставов. О болевых ощущениях пациенты сообщали редко.
Удаление коры
Когда часть коры в области SI человека повреждена или удалена хирургическим путем, возникает дефицит восприятия. Стимуляция кожи еще может быть воспринята как таковая, но способность различать пространственные детали раздражителя снижена. Степень дефицита зависит от размеров поврежденного участка коры и с течением времени постепенно уменьшается. Как полагают, это улучшение наступает вследствие способности других корковых областей принимать на себя функции.
Корреляции между соматосенсорной нейрофизиологией и психофизикой
В последнее время исследовались количественное соотношение между восприятием и уровнем активации одиночных нейронов или популяций. Полученные результаты часто позволяют делать выводы о нейронном субстрате восприятия.
В одном из опытов был применен электрический вибратор, вызывавший на кисти обезьяны синусоидальную механическую деформацию кожи разной частоты и амплитуды. Порог обнаружения (восприятия) этого стимула определяли по инструментальному условному рефлексу. Предварительно обезьянам вживляли под наркозом микроэлектрод, позволяющий одновременно регистрировать электрофизиологическую активность нейронов в SI, в частности измерять порог в реакции на механический стимул. Была установлена количественная корреляция между обоими порогами. Один из выводов, какой можно извлечь из этого опыта: данные нейроны SI участвуют в процессах восприятия действующих на кожу механических стимулов.
2.12 Контроль афферентного входа в соматосенсорной системе
На всех уровнях ЦНС афферентная информация может испытывать различные влияния и изменяться (модулироваться). Это вызывается либо взаимным торможением афферентами друг друга (афферентным торможением), либо центробежным (нисходящим) торможением. Несколько примеров афферентного торможения уже рассматривались: это аутогенное и антагонистическое торможение в спинальных двигательных системах и латеральное торможение во многих сенсорных системах.
Центробежная регуляции афферентных сигналов
Сенсорные системы нельзя рассматривать как улицы с односторонним движением, по которым информация передается от периферии к коре; напротив, на промежуточную переработку такой информации влияет целый ряд (возбуждающих или тормозных) сигналов центрального происхождения, проводимых в противоположном направлении. Схема на рис. 2.7, А показывает нисходящие тормозные пути соматосенсорной системы, начинающиеся в коре и стволе мозга.
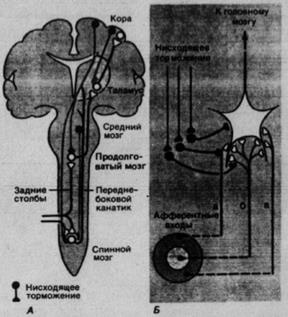
Рис. 2.7. Центробежная регуляция соматосенсорной системы. А. Афферентная информация может модулироваться в синапсах нисходящим торможением. Б. Пример функций нисходящего торможения. В синапсах, через которые активность афферентов (а, б, в) передается центральному соматосенсорному нейрону, оно может изменять величину рецептивного поля этого нейрона, если афференты, идущие от периферической части рецептивного поля (а, в), тормозятся сильнее, чем идущие от его центра (б)
Что обеспечивают эти системы нисходящего торможения? Их известные или предполагаемые функциональные влияния на афферентную информацию можно обобщить следующим образом:
- порог афферентной синаптической передачи повышается. Одно из следствий этого – подавление обыденной информации (например, непрерывной стимуляции одеждой);
- размеры рецептивного поля центрального нейрона уменьшаются при усилении нисходящего торможения (рис. 2.7, Б);
- изменяется модальность нейрона, на котором конвергируют разные типы афферентов (рис. 2.7, Б, а, б);
- регулируется чувствительность или устанавливаются пределы передачи афферентной информации.
Регуляция чувствительности афферентной передачи нисходящим торможением
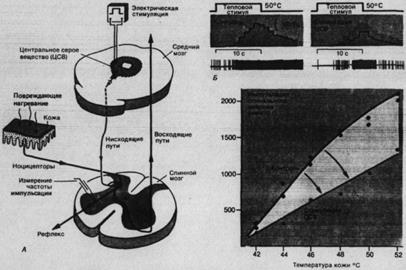
Рис. 2.8. Торможение соматосенсорной информации в спинном мозгу стимуляцией среднего мозга. А. Схема нисходящей системы и экспериментальной методики ее изучения. Стимулирующий электрод вживлен в центральное серое вещество (ЦСВ) среднего мозга наркотизированного животного. Другой электрод регистрирует здесь активность одиночных нейронов в ответ, например, на раздражение кожи очень сильным нагреванием. Б. Peaкция спинального нейрона на нагревание кожи до (слева) и во время (справа) электрической стимуляции ЦСВ. Зависимость между частотой импульсации спинального нейрона (ось ординат) и интенсивностью нагревав (температура кожи, ось абсцисс) до и во время стимуляции ЦСВ.
Рис. 2.8 показывает, что афферентная передача от кожных рецепторов может тормозиться в нейронах спинного мозга электрической стимуляцией среднего мозга. Тормозные стимулы идут по нисходящему пути и действуют либо пре-, либо постсинаптически. Тормозятся как ноцицептивные, так и неноцицептивные сигналы. В этом примере сенсорным
8-09-2015, 22:06