Аспарагин синтезируется из аспарагиновой кислоты и глутамина, который служит донором амидной группы; реакцию катализирует аспарагинсинтетаза пролин образуется из глутаминовой кислоты. Гистидин (частично заменимая аминокислота) синтезируется из АТФ и рибозы: пуриновая часть АТФ поставляет фрагмент —N=CH—NH— для имидазольного цикла гистидина; остальная часть молекулы образуется за счет рибозы.
Если в пище нет заменимой аминокислоты, клетки синтезируют ее из других веществ, и тем самым поддерживается полный набор аминокислот, необходимый для синтеза белков. Если же отсутствует хотя бы одна из незаменимых аминокислот, то прекращается синтез белков. Это объясняется тем, что в состав подавляющего большинства белков входят все 20 аминокислот; следовательно, если нет хотя бы одной из них, синтез белков невозможен.
Частично заменимые аминокислоты синтезируются в организме, однако скорость их синтеза недостаточна для обеспечения всей потребности организма в этих аминокислотах, особенно у детей. Условно заменимые аминокислоты могут синтезироваться из незаменимых: цистеин — из метионина, тирозин — из фенилаланина. Иначе говоря, цистеин и тирозин — это заменимые аминокислоты при условии достаточного поступления с пищей метионина и фенилаланина [5].
1.1.4 Участие печени в обмене витаминов
Участие печени в обмене витаминов складывается из процессов депонирования всех жирорастворимых витаминов: А, Д, Е, К, F (секреция желчи так же обеспечивает всасывание этих витаминов) и многих из гидровитаминов (В12 , фолиевая кислота, В1 , В6 , РР и др.), синтеза некоторых витаминов (никотиновая кислота) и коферментов.
Особая печени заключается в том, что в ней происходит активация витаминов:
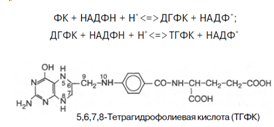
7. Фолиевая кислота с помощью витамина С восстанавливается в тетрагидрофолиевую кислоту (ТГФК) [8]; Восстановление сводится к разрыву двух двойных связей и присоединению четырех водородных атомов в положениях 5, 6, 7 и 8 с образованием тетрагидрофолиевой кислоты (ТГФК). Оно протекает в 2 стадии тканях при участии специфических ферментов, содержащих восстановленный НАДФ. Сначала при действии фолатредуктазы образуется дигидрофолиевая кислота (ДГФК), которая при участии второго фермента – дигидрофолатредуктазы – восстанавливается в ТГФК:
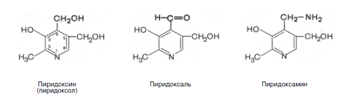
8. Витамины В1 и В6 фосфорилируются в тиаминдифосфат и пиридоксальфосфат соответственно [8]. Витамин В6 (пиридоксин) производный 3-оксипиридина. Термином витамин В6 обозначают все три производных 3-оксипиридина, обладающих одинаковой витаминной активностью: пиридоксин (пиридоксол), пиридоксаль и пиридоксамин:
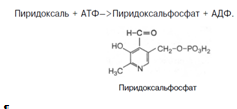
Хотя все три производных 3-оксипиридина наделены витаминными свойствами, коферментные функции выполняют только фосфорилированные производные пиридоксаля и пиридоксамина. Фосфорилирование пиридоксаля и пиридоксамина является ферментативной реакцией, протекающей при участии специфических киназ. Синтез пиридоксальфосфата, например, катализирует пиридоксалькиназа:
Витамин В1 (тиамин). В химической структуре его содержатся два кольца – пиримидиновое и тиазоловое, соединенных метиленовой связью. Обе кольцевые системы синтезируются отдельно в виде фосфорилированных форм, затем объединяются через четвертичный атом азота.
В превращении витамина B1 в его активную форму – тиаминпирофосфат (ТПФ), называемый также тиаминдифосфатом (ТДФ), участвует специфический АТФ-зависимый фермент тиаминпирофосфокиназа.
9. Часть каротинов преобразуется в витамин А под влиянием каротиндиоксигеназы. Каротины являются провитаминами для витамина А. Известны 3 типа каротинов: б-, в- и г-каротины, отличающиеся друг от друга химическим строением и биологической активностью. Наибольшей биологической активностью обладает в-каротин, поскольку он содержит два в-иононовых кольца и при распаде в организме из него образуются две молекулы витамина А:
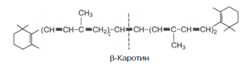
При окислительном распаде б- и г-каротинов образуется только по одной молекуле витамина А, поскольку эти провитамины содержат по одному в-иононовому кольцу.
4. Витамин Д подвергается первому гидроксилированию на пути получения гормона кальцитриола; в печени осуществляется гидроксилирование в 25-м положении. Ферменты, катализирующие эти реакции, называются гидроксилазами, или монооксигеназами. В реакциях гидроксилирования используется молекулярный кислород.
5. Окислившийся витамин С восстанавливается в аскорбиновую кислоту;
6. Витамины РР, В2 , пантотеновая кислота включаются в соответствующие нуклеотиды (НАД+ , НАД+ Ф, ФМН, ФАД, КоА-SH);
7. Витамин К окисляется, чтобы в виде своего пероксида служить коферментом в созревании (посттрансляционной модификации) белковых факторов свёртывания крови.
В печени синтезируются белки, выполняющие транспортные функции по отношению к витаминам. Например, ретинолсвязывающий белок (его содержание уменьшается при опухолях), витамин Е-связывающий белок и т.д. Часть витаминов, в первую очередь жирорастворимых, а также продуктов их преобразований выделяется из организма в составе жёлчи.
1.1.5 Участие печени в водно-минеральном обмене
Участие печени в водно-минеральном обмене состоит в том, что она дополняет деятельность почек в поддержании водно-солевого равновесия и является как бы внутренним фильтром, организма. Печень задерживает ионы Na+ , К+ , Сl- , Ca2+ и воду и выделяет их в кровь. Кроме того, печень депонирует макро- (К, Na, Ca, Mg, Fe) и микро- (Cu, Mn, Zn, Cо, As, Cd, Pb, Se) элементы и участвует в их распределении по другим тканям с помощью транспортных белков.
Для накопления железа гепатоциты синтезируют специальный белок – ферритин. В ретикулоэндотелиоцитах печени и селезенки регистрируется водонерастворимый железосодержащий протеиновый комплекс – гемосидерин. В гепатоцитах синтезируется церулоплазмин, который, кроме вышеназванных функций, выполняет роль транспортного белка для ионов меди. Трансферрин, обладающий как и церулоплазмин, полифункциональностью, также образуется в печени и используется для переноса в плазме крови только ионов железа. Данный белок необходим для эмбрионального клеточного роста в период формирования печени. В печени ион Zn включается в алкогольдегидрогеназу, необходимую для биотрансформации этанола. Поступившие в гепатоциты соединения селена преобразуются в Se-содержащие аминокислоты и с помощью специфической т-РНК включаются в различные Se-протеины: глутатионпероксидазу (ГПО), 1-йодтиронин-5’- дейодиназу, Se-протеин Р. Последний считают основным транспортёром этого микроэлемента. Дейодиназа, обнаруженная не только в печени, обеспечивает конверсию прогормона тироксина в активную форму – трийодтиронин. Как известно, глутатионпероксидаза – ключевой фермент антирадикальной защиты. В печени сера, включённая в аминокислоты, окисляется до сульфатов, которые в виде ФАФС (фосфоаденозилфосфосульфатов) используются в реакциях сульфирования ГАГов, липидов, а также в процессах биотрансформации ксенобиотиков и некоторых эндогенных веществ (примеры продуктов инактивации – скатоксилсульфат, индоксилсульфат). Печень способна служить временным депо воды, особенно при отёках (количество Н2 О может составлять до 80 % от массы органа) [6].
1.1.6 Участие печени в пигментном обмене
Участие печени в обмене пигментов проявляется в превращении хромопротеидов до билирубина в клеткахРЭС, имеющихся в печени, конъюгации билирубина в самих печеночных клетках и разложении в них всасывающегося из кишечника уробилиногена до непигментных продуктов.
Гемохромогенные пигменты, образуются в организме при распаде гемоглобина (в значительно меньшей степени при распаде миоглобина, цитохромов и др.).
Начальным этапом распада гемоглобина (в клетках макрофагов, в частности в звездчатых ретикулоэндотелиоцитах, а также в гистиоцитах соединительной ткани любого органа) является разрыв одного метинового мостика с образованием вердоглобина. В дальнейшем от молекулы вердоглобина отщепляются атом железа и белок глобин. В результате образуется биливердин, который представляет собой цепочку из четырех пиррольных колец, связанных метановыми мостиками. Затем биливердин, восстанавливаясь, превращается в билирубин – пигмент, выделяемый с желчью и поэтому называемый желчным пигментом. Образовавшийся билирубин называется непрямым (неконъюгированным) билирубином. Он нерастворим в воде, дает непрямую реакцию с диазореактивом, т.е. реакция протекает только после предварительной обработки спиртом. В печени билирубин соединяется (конъюгирует) с глюкуроновой кислотой. Эта реакция катализируется ферментом УДФ-глюкуронилтрансферазой, при этом глюкуроновая кислота вступает в реакцию в активной форме, т.е. в виде УДФГК. Образующийся глюкуронид билирубина получил название прямого билирубина (конъюгированный билирубин). Он растворим в воде и дает прямую реакцию с диазореактивом. Большая часть билирубина соединяется с двумя молекулами глюкуроновой кислоты, образуя диглюкуронид билирубина. Образовавшийся в печени прямой билирубин вместе с очень небольшой частью непрямого билирубина выводится с желчью в тонкую кишку. Здесь от прямого билирубина отщепляется глюкуроновая кислота и происходит его восстановление с последовательным образованием мезобилирубина и мезобилиногена (уробилиногена). Из тонкой кишки часть образовавшегося мезобилиногена (уробилиногена) резорбируется через кишечную стенку, попадает в воротную вену и током крови переносится в печень, где расщепляется полностью до ди- и трипирролов. Таким образом, в норме в общий круг кровообращения и мочу мезобилиноген не попадает. Основное количество мезобилиногена из тонкой кишки поступает в толстую и здесь восстанавливается до стеркобилиногена при участии анаэробной микрофлоры. Образовавшийся стеркобилиноген в нижних отделах толстой кишки (в основном в прямой кишке) окисляется до стеркобилина и выделяется с калом. Лишь небольшая часть стеркобилиногена всасывается в систему нижней полой вены (попадает сначала в геморроидальные вены) и в дальнейшем выводится с мочой (приложение 18).
1.2 Мочевинообразовательная функция
Печень — единственный орган, имеющий все ферменты цикла образования мочевины из аммиака. Аммиак, образующийся в других тканях, в печени превращается в индифферентный продукт— мочевину, которая выделяется в кровь. При интенсивном катаболизме белков и небелковых азотистых соединений (аминокислот, пуринов, пиримидинов, биогенных аминов) повышено образование мочевины в печени, ее содержание в крови и выделение с мочой.
Биосинтез мочевины (орнитиновый цикл мочевинообразования) является основным механизмом обезвреживания аммиака в организме (приложение 19). На долю мочевины приходится до 80–85% от всего азота мочи. Впервые Г. Кребс и К. Гензеляйт в 1932 г. вывели уравнения реакций синтеза мочевины, которые представлены в виде цикла, получившего в литературе название орнитинового цикла мочевинообразования Кребса.
Весь цикл мочевинообразования может быть представлен следующим образом. На первом этапе синтезируется макроэргическое соединение карбамоилфосфат – метаболически активная форма аммиака, используемая в качестве исходного продукта для синтеза пиримидиновых нуклеотидов (соответственно ДНК и РНК) и аргинина (соответственно белка и мочевины):
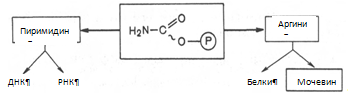
Существуют два разных пути синтеза карбамоилфосфата в клетках печени, катализируемые разными ферментами. Первую необратимую реакцию катализирует регуляторный фермент – аммиакзависимая карбамоилфосфатсинтетаза:
![]()
Реакция требует затраты двух молекул АТФ, проходит в митохондриях клеток печени и используется преимущественно для синтеза аргинина и мочевины. В этой реакции в качестве активного стимулирующего аллостерического эффектора действует N-ацетилглутамат. Вторую, также необратимую, реакцию катализирует глутаминзависимая карбамоилфосфатсинтетаза:
![]()
Данная реакция открыта в цитозоле клеток печени и требует наличия ионов Mg2+ .
На втором этапе цикла мочевинообразования происходит конденсация карбамоилфосфата и орнитина с образованием цитруллина; реакцию катализирует орнитин-карбамоилтрансфераза. На следующей стадии цитруллин превращается в аргинин в результате двух последовательно протекающих реакций. Первая из них, энергозависимая,– это конденсация цитруллина и аспарагиновой кислоты с образованием аргининосукцината (эту реакцию катализирует аргининосукцинатсинтетаза). Аргининосукцинат распадается в следующей реакции на аргинин и фумарат при участии другого фермента – аргининосукцинатлиазы. На последнем этапе аргинин расщепляется на мочевину и орнитин под действием аргиназы. Суммарная реакция синтеза мочевины без учета всех промежуточных продуктов может быть представлена в следующем виде:
![]()
Учитывая выше изложенные данные о механизмах обезвреживания аммиака в организме, можно сделать следующее заключение. Часть амиака используется на биосинтез аминокислот путем восстановительного аминирования б-кетокислот по механизму реакции трансаминирования. Аммиак связывается при биосинтезе глутамина и аспарагина. Некоторое количество аммиака выводится с мочой в виде аммонийных солей. В форме креатинина, который образуется из креатина и креатинфосфата, выделяется из организма значительная часть азота аминокислот. Наибольшее количество аммиака расходуется на синтез мочевины, которая выводится с мочой в качестве главного конечного продукта белкового обмена в организме человека и животных. Подсчитано, что в состоянии азотистого равновесия организм взрослого здорового человека потребляет и соответственно выделяет примерно 15 г азота в сутки; из экскретируемого с мочой количества азота на долю мочевины приходится около 85%, креатинина – около 5%, аммонийных солей – 3%, мочевой кислоты – 1% и на другие формы – около 6% [2].
1.3 Желчеобразовательная и экскреторная функция
Печень образует специальный жидкий экскрет — желчь, которая выделяется в тонкий кишечник. Нарушение экскреторной функции неблагоприятно влияет на переваривание и всасывание липидов и вызывает накопление токсических продуктов обмена пигментов и чужеродных веществ. Компонентами жёлчи, кроме воды, являются соли жёлчных кислот, холестерин, фосфолипиды, билирубин-глюкурониды, муцин, немного белков (альбумины, 5’-нуклеотидаза, щелочная фосфатаза, ГГТП, иммуноглобулин А), различных ионов (Na+ , Cl- , HCO3 - , Cu++ , Fe+++ , Se++ и др.), микроколичества липовитаминов, стероидных гормонов и т.д.
Желчь – жидкий секрет желтовато-коричневого цвета, в сутки у человека образуется 500–700 мл желчи (10 мл на 1 кг массы тела). Желчеобразование происходит непрерывно, хотя интенсивность этого процесса на протяжении суток резко колеблется. Вне пищеварения печеночная желчь переходит в желчный пузырь, где происходит ее сгущение в результате всасывания воды и электролитов. Относительная плотность печеночной желчи 1,01, а пузырной – 1,04. Концентрация основных компонентов в пузырной желчи в 5–10 раз выше, чем в печеночной.
Образование желчи начинается с активной секреции гепатоцитами воды, желчных кислот и билирубина, в результате которой в желчных канальцах появляется так называемая первичная желчь. Она, проходя по желчным ходам, вступает в контакт с плазмой крови, вследствие чего между желчью и плазмой устанавливается равновесие электролитов, т.е. в образовании желчи принимают участие в основном два механизма – фильтрация и секреция.
В печеночной желчи можно выделить две группы веществ. Первая группа – это вещества, которые присутствуют в желчи в количествах, мало отличающихся от их концентрации в плазме крови (например, ионы Na, К, креатин и др.), что в какой-то мере служит доказательством наличия фильтрационного механизма. Ко второй группе относятся соединения, концентрация которых в печеночной желчи во много раз превышает их содержание в плазме крови (билирубин, желчные кислоты и др.), что свидетельствует о наличии секреторного механизма. В последнее время появляется все больше данных о преимущественной роли активной секреции в механизме желчеобразования. Кроме того, в желчи обнаружен ряд ферментов, из которых особо следует отметить щелочную фосфатазу печеночного происхождения. При нарушении оттока желчи активность данного фермента в сыворотке крови возрастает.
Основные функции желчи. 1. Эмульсификация. Соли желчных кислот обладают способностью значительно уменьшать поверхностное натяжение. Благодаря этому они осуществляют эмульгирование жиров в кишечнике, растворяют жирные кислоты и нерастворимые в воде мыла. 2. Нейтрализация кислоты. Желчь, рН которой немногим более 7,0, нейтрализует кислый химус, поступающий из желудка, подготавливая его для переваривания в кишечнике. 3. Экскреция. Желчь – важный носитель экскретируемых желчных кислот и холестерина. Кроме того, она удаляет из организма многие лекарственные вещества, токсины, желчные пигменты и различные неорганические вещества, такие, как медь, цинк и ртуть. 4. Растворение холестерина. Как отмечалось, холестерин, подобно высшим жирным кислотам, представляет собой нерастворимое в воде соединение, которое сохраняется в желчи в растворенном состоянии лишь благодаря присутствию в ней солей желчных кислот и фосфатидилхолина. При недостатке желчных кислот холестерин выпадает в осадок, при этом могут образовываться камни. Обычно камни имеют окрашенное желчным пигментом внутреннее ядро, состоящее из белка. Чаще всего встречаются камни, у которых ядро окружено чередующимися слоями холестерина и билирубината кальция. Такие камни содержат до 80% холестерина. Интенсивное образование камней отмечается при застое желчи и наличии инфекции. При застое желчи встречаются камни, содержащие 90–95% холестерина, а при инфекции могут образовываться камни, состоящие из билирубината кальция. Принято считать, что присутствие бактерий сопровождается увеличением в-глюкуронидазной активности желчи, что приводит к расщеплению конъюгатов билирубина; освобождающийся билирубин служит субстратом для образования камней.
1.4 Биотрансформационная (обезвреживающая) функция
Печень является основным органом, где происходит обезвреживание природных продуктов обмена и чужеродных веществ. Чужеродные вещества (ксенобиотики) в печени превращаются в менее токсичные и индифферентные вещества. В этом смысле можно говорить об обезвреживании, но более точно применяется термин биотрансформация. Эти процессы проходят путем окисления, восстановления, метилирования, ацетилирования и конъюгации с теми или иными веществами. Окисление, восстановление и гидролиз чужеродных соединений осуществляют в основном микросомальные ферменты. Наряду с микросомальным в печени существует также пероксисомальное окисление.
Пероксисомы – микротельца, обнаруженные в гепатоцитах; их можно рассматривать как специализированные окислительные органеллы. Эти микротельца содержат оксидазу мочевой кислоты, лактатоксидазу, оксидазу D-аминокислот, а также каталазу (катализирует расщепление перекиси водорода, которая образуется при действии указанных оксидаз). Пероксисомальное окисление, так же как и микросомальное, не сопровождается образованием макроэргических связей.
В печени широко представлены также защитные синтезы, например синтез мочевины, в результате которого обезвреживается весьма токсичный аммиак. Это было рассмотренно в мочевинообразавательной функции. В результате гнилостных процессов, протекающих в кишечнике, из тирозина образуются фенол и крезол, а из триптофона – скатол и индол. Эти вещества всасываются и с током крови поступают в печень, где обезвреживаются путем образования парных соединений с серной или глюкуроновой кислотой. Обезвреживание фенола, крезола, скатола и индола в печени происходит в результате взаимодействия этих соединений не со свободными серной и глюкуроновой кислотами, а с их так называемыми активными формами: ФАФС и УДФГК. Глюкуроновая кислота участвует не только в обезвреживании продуктов гниения белковых веществ, образовавшихся в кишечнике, но и в связывании ряда других токсичных соединений, образующихся в процессе обмена в тканях. В частности, свободный, или непрямой, билирубин, обладающий значительной токсичностью, в печени взаимодействует с глюкуроновой кислотой, образуя моно- и диглюкурониды билирубина. Нормальным метаболитом является и гиппуровая кислота, образующаяся в печени из бензойной кислоты и глицина. Синтез гиппуровой кислоты у человека протекает преимущественно в печени.
В печени широко представлены процессы метилирования. Так, перед выделением с мочой амид никотиновой кислоты (витамин РР) метилируется
8-09-2015, 20:08