Внутренняя общая система костных пластинок граничит с эндоостом костной полоси и представлена пластинками, ориентированными параллельно поверхности канала.
Различают два вида остеогенеза: непосредственно из мезенхимы («прямой») и путем замещения костью эмбрионального хряща («непрямой») остеогенез – рис. 19,20.
Первый характерен для развития грубоволокнистой кости черепа и нижней челюсти. Процесс начинается с интенсивного развития соединительной ткани и кровеносных сосудов. Мезенхимные клетки, анастомозируя между собой отростками, образуют сеть. Клетки, оттесненные межклеточным веществом на поверхность, дифференцируются в остеобласты, активно участвующие в остеогенезе. В последующем первичная грубоволокнистая костная ткань замещается пластинчатой костью. Кости туловища, конечностей и др. формируются на месте хрящевой ткани. В трубчатых костях этот процесс начинается в области диафиза формированием под надхрящницей сети перекладин грубоволокнистой кости - костной манжетки. Процесс замещения хряща костной тканью называется энхондральным окостенением.
Одновременно с развитием энхондральной кости со стороны надкостницы идет активный процесс перихондрального остеогенеза, формирующий плотный слой периостальной кости, распространяющийся по всей ее длине до эпифизарной пластинки роста. Периостальная кость представляет компактное вещество кости скелета.
Позднее центры окостенения появляются в эпифизах кости. Костная ткань здесь замещает хрящевую. Последняя сохраняется лишь на суставной поверхности и в эпифизарной пластинке роста, отграничивающей эпифиз от диафиза в течение всего периода роста организма до половой зрелости животного.
Надкостница (периост) состоит из двух слоев: внутренний - содержит коллагеновые и эластические волокна, остеобластыЮ остеокласты и кровеносные сосуды. Наружный - образован плотной соединительной тканью. Она непосредственно связана с сухожилиями мышц.
Эндоост - слой соединительной ткани, выстилающий костно-мозговой канал. Он содержит остеобласты и тонкие пучки коллагеновых волокон, переходящих в ткань костного мозга.
Мышечные ткани
1. Гладкие.
2. Сердечная поперечнополосатая.
3. Скелетные поперечнополосатые.
4. Развитие, рост и регенерация мышечных волокон.
1. Ведущая функция мышечных тканей - обеспечение перемещения в пространстве организма в целом и его частей. Все мышечные ткани составляют морфофункциональную группу, а в зависимости от структуры органелл сокращения ее делят на три группы: гладкие, скелетные поперечно-полосатые и сердечные поперечнополосатые мышечные ткани. Единого источника эмбрионального развития у этих тканей нет. Ими являются мезенхима, миотомы сегментированной мезодермы, висцеральный листок спланхнотома и др.
Гладкие мышечные ткани мезенхимного происхождения. Ткань состоит из миоцитов и соединительнотканного компонента. Гладкий миоцит представляет собой веретеновидную клетку длиной 20-500 мкм, толщиной 5-8 мкм. Ядро палочковидной формы находится в ее центральной части. В клетке много митохондрий.
Каждый миоцит окружен базальной мембраной. В ней есть отверстия, в области которых между соседними миоцитами образуются щелевидные соединения (нексусы), обеспечивающие функциональные взаимодействия миоцитов в ткани. В базальную мембрану вплетены многочисленные ретикулярные фибриллы. Вокруг мышечных клеток ретикулярные, эластические и тонкие коллагеновые волокна образуют трехмерную сеть - эндомизий, которая объединяет соседние миоциты.
Физиологическая регенерация гладкой мышечной ткани проявляется обычно в условиях повышенных функциональных нагрузок преимущественно в форме компенсаторной гипертрофии. Наиболее отчетливо это наблюдается в мышечной оболочке матки во время беременности.
Элементами мышечной ткани эпидермального происхождения являются миоэпителиальные клетки, развивающиеся из эктодермы. Они располагаются в потовых, молочных, слюнных и слезных железах, дифференцируясь одновременно с их секреторными эпителиальными клетками из общих предшественников. Сокращаясь, клетки способствуют выведению секрета железы.
Гладкие мышцы образуют мышечные слои во всех полых и трубчатых органах.
2. Источники развития сердечной поперечнополосатой мышечной ткани - симметричные участки висцерального листка спланхнотома. Большинство ее клеток дифференцируются в кардиомиоциты (сердечные миоциты), остальные - в клетки мезотелия эпикарда. И те и другие имеют общие клетки-предшественники. В ходе гистогенеза дифференцируются несколько видов кардиомиоцитов: сократительные, проводящие, переходные и секреторные.
Строение сократительных кардиомиоцитов. Клетки имеют удлинненную форму (100-150 мкм), близкую к цилиндрической. Их концы соединяются друг с другом вставочными дисками. Последние выполняют не только механическую функцию, но и проводящую, обеспечивают электрическую связь между клетками. Ядро овальной формы, располагается в центральной части клетки. В ней много митохондрий. Они образуют цепочки вокруг специальных органелл - миофибрилл. Последние построены из постоянно существующих упорядоченно расположенных нитей актина и миозина - сократительных белков. Для их закрепления служат особые структуры - телофрагма и мезофрагма, построенные из других белков.
Участок миофибриллы между двумя Z -линиями называется саркомером. А-полосы - анизотропные, микрофиламенты толстые, содержат миозин: I-полосы - изотропные, микрофиламенты тонкие, содержат актин; H-полоса располагается посредине А-полосы (рис.21).
Существует несколько теорий механизма сокращения миоцитов:
1) Под влиянием потенциала действия, который распространяется по цитолемме, ионы кальция освобождаются, поступают к миофибриллам и инициируют сократительный акт, являющийся результатом взаимодействия актиновых и миозиновых микрофиламентов; 2) Наиболее распространенной в настоящее время теорией является модель скользящих нитей (Г. Хаксли, 1954). Мы являемся сторонниками последней.
Особенности строения проводящих кардиомиоцитов. Клетки крупнее рабочих кардиомиоцитов (длина около 100 мкм, а толщина около 50 мкм). Цитоплазма содержит все органеллы общего значения. Миофибриллы немногочисленны и лежат по периферии клетки. Эти кардиомиоциты соединяются в волокна друг с другом не только концами, но и боковыми поверхностями. Основная функция проводящих кардиомиоцитов состоит в том, что они воспринимают управляющие сигналы от пейсмекерных элементов и передают информацию к сократительным кардиомиоцитам (рис.22).
В дефинитивном состоянии сердечная мышечная ткань не сохраняет ни стволовых клеток, ни клеток-предшественников, поэтому, если кардиомиоциты гибнут (инфаркт), то они не восстанавливаются.
3. Источником развития элементов скелетной поперечнополосатой мышечной ткани являются клетки миоциты. Одни из них дифференцируются на месте, другие же мигрируют из миотомов в мезенхиму. Первые участвуют в формировании миосимпласта, вторые дифференцируются в миосателлитоциты.
Основным элементом скелетной мышечной ткани является мышечное волокно, образованное миосимпластом и миосателлитоцитами. Волокно окружено сарколеммой. Поскольку симпласт не клетка, термин “цитоплазма” не применяют, а говорят “саркоплазма” (греч. sarcos - мясо). В саркоплазме у полюсов ядер располагаются органеллы общего значения. Специальные органеллы представлены миофибриллами.
Механизм сокращения волокон такой же, как и в кардиомиоцитах.
Большую роль в деятельности мышечных волокон играют включения, в первую очередь миоглобина и гликогена. Гликоген служит основным источником энергии, необходимой как для совершения мышечной работы, так и для поддержания теплового баланса всего организма.
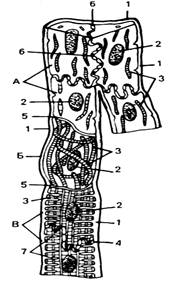
Рис. 22. Ультрамикроскопическое строение трех видов кардиомиоцитов: проводящих (А), промежуточных (Б) и рабочих (В) (схема по Г.С. Катинасу)
1 — базальная мембрана; 2 — ядра клеток; 3 — миофибриллы; 4 — плазмолемма; 5 — соединение рабочих кардиомиоцитов (вставочный диск); соединения промежуточного кардиомиоцита с рабочим и проводящим кардиомиоцитами; 6 — соединение проводящих кардиомиоцитов; 7 — поперечные трубочки-системы (органеллы общего назначения не показаны).
Миосателлитоциты прилежат к поверхности симпласта так, что их плазмолеммы соприкасаются. С одним симпластом связано значительное количество сателлитоцитов. Каждый миосателлитоцит - одноядерная клетка. Ядро мельче, чем ядро миосимпласта, и более округлое. Митохондрии и эндоплазматическая сеть распределены в цитоплазме равномерно, комплекс Гольджи и клеточный центр расположены рядом с ядром. Миосателлитоциты - камбиальные элементы скелетной мышечной ткани.
Мышца как орган. Между мышечными волокнами находятся тонкие прослойки рыхлой соединительной ткани - эндомизий. Его ретикулярные и коллагеновые волокна переплетаются с волокнами сарколеммы, что способствует объединению усилий при сокращении. Мышечные волокна группируются в пучки, между которыми располагаются более толстые прослойки рыхлой соединительной ткани - перимизий. В нем содержатся также и эластические волокна. Соединительная ткань, окружающая мышцу в целом, называется эпимизием.
Васкуляризация. Артерии, вступающие в мышцу, ветвятся в перимизии. Рядом с ними много тканевых базофилов, регулирующих проницаемость сосудистой стенки. Капилляры располагаются в эндомизии. Венулы и вены лежат в перимизии рядом с артериолами и артериями. Здесь же проходят и лимфососуды.
Иннервация. Нервы, вступающие в мышцу содержат как эфферентные, так и афферентные волокна. Отросток нервной клетки, приносящий эфферентный нервный импульс, проникает через базальную мембрану и ветвится между ней и плазмолеммой симпласта, участвуя в образовании двигательной, или моторной бляшки. Нервный импульс освобождает здесь медиаторы, которые вызывают возбуждение, распространяющееся по плазмолемме симпласта.
Итак, каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров. Этот комплекс образует морфофункциональную единицу скелетной мышцы - мион; иногда мионом называют само мышечное волокно, что не соответствует Международной гистологической номенклатуре.
4. Клетки, из которых в эмбриогенезе образуются поперечнополосатые мышечные волокна, называются миобластами. После ряда делений эти одноядерные клетки, не содержащие миофибрилл, начинают сливаться между собой, формируя удлиненные многоядерные цилиндрические образования - микротрубочки, в которых в свое время появляются миофибриллы и другие органеллы, характерные для поперечнополосатых мышечных волокон. У млекопитающих большинство этих волокон образуется еще до рождения. В период постнатального роста мышцы должны становиться длиннее и толще для того, чтобы сохранить соразмерность с растущим скелетом. Их окончательная величина зависит от выпадающей на их долю работы. После первого года жизни дальнейший рост мышц всецело обусловлен утолщением отдельных волокон, т. е. представляет собой гипертрофию (гипер - над, сверх и трофи - питание), а не увеличением их числа, что называлось бы гиперплазией (от плазис - образование).
Таким образом, поперечнополосатые мышечные волокна растут в толщину путем увеличения числа содержащихся в них миофибрилл ( и других органелл).
Мышечные волокна удлиняются в результате слияния с клетками-сателлитами. Кроме того, в постнатальном периоде возможно удлинение миофибрилл путем пристройки к их концам новых саркомеров.
Регенерация. Клетки-сателлиты не только обеспечивают один из механизмов роста поперечнополосатых мышечных волокон, но и остаются в течение всей жизни потенциальным источником новых миобластов, слияние которых может приводить к образованию совершенно новых мышечных волокон. Клетки-сателлиты способны делиться и давать начало миобластам после мышечной травмы и при некоторых дистрофических состояниях, когда наблюдаются попытки регенерации новых волокон. Однако даже незначительные дефекты мышечной ткани после тяжелых травм заполняются фиброзной тканью, образуемой фибробластами.
Рост и регенерация гладких мышц. Подобно другим типам мышц, гладкие мышцы отвечают на повышенные функциональные требования компенсаторной гипертрофией, но это не единственная возможная реакция. Например, во время беременности увеличиваются не только размеры гладких мышечных клеток в стенке матки (гипертрофия), но и их число (гиперплазия).
У животных при беременности или после введения гормонов в мышечных клетках матки часто можно видеть фигуры митоза; поэтому общепризнано, что гладкие мышечные клетки сохраняют способность к митотическому делению.
Нервная ткань
1. Развитие ткани.
2. Классификация нервных клеток.
3. Нейроглия, её разновидность.
4. Синапсы, волокна, нервные окончания.
1. Нервная ткань - специализированная ткань, которая формирует основную интегрирующую систему организма - нервную систему. Основная функция - проводимость.
Нервная ткань состоит из нервных клеток - нейронов, выполняющих функцию нервного возбуждения и проведения нервного импульса, и нейроглии, обеспечивающей опорную, трофическую и защитную функции.
Нервная ткань развивается из дорсального утолщения эктодермы - нервной пластинки, которая в процессе развития дифференцируется в нервную трубку, нейральные гребни (валики) и нейральные плакоды.
В последующие периоды эмбриогенеза из нервной трубки образуется головной и спинной мозг. Нейральный гребень формирует чувствительные ганглии, ганглии симпатической нервной системы, меланоциты кожи и т. д. Нейральные плакоды участвуют в формировании органов обоняния, слуха, чувствительных ганглиев.
Нервная трубка состоит из одного слоя призматических клеток. Последние, размножаясь, образуют три слоя: внутренний - эпендимный, средний - мантийный и наружный - краевую вуаль.
В последующем клетки внутреннего слоя продуцируют эпендимные клетки, выстилающие центральный канал спинного мозга. Клетки мантийного слоя дифференцируются на нейробласты, превращающиеся далее в нейроны и спонгиобласты, дающие начало различным видам нейроглии (астроцитам, олигодендроцитам).
2. Нервные клетки (нейроциты, нейроны) различных отделов нервной системы характеризуются разнообразием формы, размеров и функционального значения. В соответствии с функцией нервные клетки делятся на рецепторные (афферентные), ассоциативные и эффекторные (эфферентные).
При большом разнообразии формы нервных клеток общим морфологическим признаком является наличие отростков, обеспечивающих их связь в составе рефлекторных дуг. Длина отростков различна и колеблется от нескольких микрон до 1-1,5 м.
Отростки нервных клеток по функциональному значению делят на два вида. Одни принимают нервное возбуждение и проводят его к перикариону нейрона. Они получили название дендритов. Другой вид отростков проводит импульс от тела клетки и передает его на другой нейроцит или на аксон (axos - ось), или нейрит. Все нервные клетки имеют только один нейрит.
По количеству отростков нервные клетки делят на униполярные - с одним отростком, биполярные и мультиполярные (рис.23).
Ядра нервных клеток крупные, округлые или слегка овальные, располагаются в центре перикариона.
Цитоплазма клеток характеризуется обилием различных органелл, нейрофибрилл, хроматофильного вещества. Поверхность клетки покрыта плазмолеммой, которая характеризуется возбудимостью и способностью проводить возбуждение.
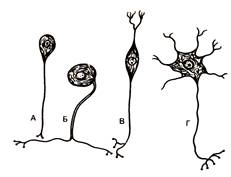
Рис. 23. Типы нервных клеток (схема по Т.Н. Радостиной, Л.С. Румянцевой)
А – униполярный нейрон; Б - псевдоуниполярный нейрон; В – биполярный нейрон; Г – мультиполярный нейрон.
Нейрофибриллы - совокупность волокон, структур цитоплазмы, формирующие в перикарионе густое сплетение.
Хроматофильное (базофильное) вещество выявляется в перикарионе нефроцитов и в их дендритах, но отсутствует в аксонах.
Эпендимоциты выстилают полости центральной нервной системы: желудочки головного мозга и спинно-мозговой канал. Клетки, обращенные в полость нервной трубки, содержат реснички. Их противоположные полюса переходят в длинные отростки, которые поддерживают остов тканей нервной трубки. Эпендимоциты участвуют в секреторной функции, выделяя в кровь различные активные вещества.
Астроциты бывают протоплазматические (коротколучистые) и волокнистые (длиннолучистые). Первые локализованы в сером веществе ЦНС ( центральной нервной системы). Они участвуют в обмене веществ нервной ткани и выполняют разграничительную функцию.
Волокнистые астроциты характерны для белого вещества ЦНС. Они образуют поддерживающий аппарат ЦНС.
Олигодендроциты - большая группа клеток ЦНС и ПНС (периферическая нервная система). Они окружают тела нейронов, входят в состав оболочек нервных волокон и нервных окончаний, участвуют в их обмене веществ.
Микроглия (глиальные макрофаги) - специализированная система макрофагов, выполняющая защитную функцию. Они развиваются из мезенхимы, способны к амебоидному перемещению. Они характерны для белого и серого вещества ЦНС.
4. Отростки нервных клеток в совокупности с покрывающими их клетка2ми нейроглии образуют нервные волокна. Расположенные в них отростки нервных клеток называют осевыми цилиндрами, а покрывающие их клетки олигодендроглии - нейролеммоцитами (шванновские клетки).
Различают миелиновые и безмиелиновые нервные волокна.
Безмиелиновые (безмякотные) нервные волокна характерны для вегетативной нервной системы. Леммоциты плотно прилегают друг к другу, образуя непрерывные тяжи. В волокне содержится несколько осевых цилиндров, т. е. отростков различных нервных клеток. Плазмолемма образует глубокие складки, формирующие сдвоенную мембрану - мезаксон, на котором подвешен осевой цилиндр. При световой микроскопии эти структуры не выявляются, что создает впечатление погружения осевых цилиндров непосредственно в цитоплазму глиальных клеток.
Миелиновые (мякотные) нервные волокна. Их диаметр колеблется от 1 до 20 мкм. Они содержат один осевой цилиндр - дендрит или нейрит нервной клетки, покрытой оболочкой, образованной леммоцитами. В оболочке волокна различают два слоя: внутренний - миелиновый, более толстый и наружный - тонкий, содержащий цитоплазму и ядра леммоцитов.
На границе двух леммоцитов оболочка миелинового волокна истончается, образуется сужение волокна - узловой перехват (перехват Ранвье). Участок нервного волокна между двумя перехватами называется межузловым сегментом. Его оболочка соответствует одному леммоциту.
Нервные окончания различны по их функциональному значению. Различают три вида нервных окончаний: эффекторные, рецепторные и концевые аппараты.
Эффекторные нервные окончания - к их числу относят двигательные нервные окончания поперечнополосатых и гладких мышц и секреторные окончания железистых органов.
Двигательные нервные окончания поперечнополосатых скелетных мышц - моторные бляшки - комплекс взаимосвязанных структур нервной и мышечной тканей.
Чувствительные нервные окончания (рецепторы) - специализированные концевые образования дендритов чувствительных нейронов. Различают две большие группы рецепторов: экстерорецепторы и интерорецепторы. Чувствительные окончания делятся на механорецепторы, хеморецепторы, терморецепторы и др. Они подразделяются на свободные нервные окончания и несвободные. Последние покрыты соединительнотканной капсулой и называются инкапсулированными. К этой группе относят пластинчатые тельца (тельца Фатера-Пачини), осязательные тельца (тельца Мейснера) и др.
Пластинчатые тельца характерны для глубоких слоев кожи и внутренних органов. Осязательные тельца также образованы клетками глии.
Синапсы - специализированный контакт двух нейронов, обеспечивающий одностороннее проведение нервного возбуждения. Морфологически в синапсе различают пресинаптический и постсинаптический полюсы, а между ними щель. Встречаются синапсы с химической и электрической передачей.
По месту контакта различают синапсы: аксосоматические, аксодендрические и аксоаксональные.
Пресинаптический полюс синапса характеризуется наличием синаптических пузырьков, содержащих медиатор (ацетилхолин или норадреналин).
Нервная система представлена чувствительными и двигательными клетками, объединенными межнейрональными синапсами в функционально активные образования - рефлекторные дуги. Простая рефлекторная дуга состоит из двух нейронов - чувствительного и двигательного.
Рефлекторные дуги высших позвоночных животных содержат в своем составе ещё значительное количество ассоциативных нейронов, расположенных между чувствительными и двигательными нейронами.
Нерв - это пучок волокон, окруженных плотной оболочкой периневрия. Мелкие нервы состоят только из одного пучка, окруженного эндоневрием. Число и диаметр нервных волокон в пучке весьма непостоянны. В дистальных участках некоторых нервов имеется больше волокон, чем в более проксимальных. Это объясняется ветвлением волокон.
Кровоснабжение нервов. Нервы обильно снабжены сосудами, образующими много анастомозов. Имеются эпиневральные, межпучковые, периневральные и внутрипучковые артерии и артериолы. Эндоневрий содержит сеть капилляров.
Литература
1. Александровская О.В., Радостина Т.Н., Козлов Н.А. Цитология, гистология и эм бриология.-М:Агропромиздат, 1987.- 448 с.
2. Афанасьев Ю.И., Юрина Н.А. Гистология.- М: Медицина, 1991.- 744 с.
3. Вракин В.Ф., Сидорова М.В. Морфология сельскохозяйственных животных. - М: Агропромиздат, 1991.- 528 с.
4. Глаголев П.А., Ипполитова В.И. Анатомия сельскохозяйственных животных с основами гистологии и эмбриологии.- М: Колос, 1977.- 480 с.
5. Хэм А., Кормак Д. Гистология. -М: Мир, 1982.-Т 1-5.
6. Серавин Л.Н. Происхождение эукариотической
8-09-2015, 23:35