Министерство образования Российской Федерации
Федеральное агентство по образованию
Государственное образовательное учреждение
Высшего профессионального образования
Дипломная работа
Тема: "Распределение антигенов системы HLAу больных туберкулезом и здоровых представителей русской этнической группы Челябинской области"
Работа допущена к защите
Заведующий кафедрой
Дата представления
Дата защиты
Оценка
Челябинск 2008
![]() Задание на выполнение дипломной работы
Задание на выполнение дипломной работы
Тема: Распределение антигенов системы HLA у больных туберкулезом и здоровых представителей русской этнической группы Челябинской области
Утверждена " " 2008 года
Срок сдачи студентом законченной работы " " 2008 года.
Перечень работ Изучить особенности распределения генов HLAII класса и их гаплотипов в группе больных туберкулезом; сделать вывод о наличии положительной или отрицательной ассоциации генов HLAII класса с туберкулезом у представителей русской этнической группы Челябинской области
Содержание дипломной работы: Выявить ассоциации генов HLAII класса с развитием туберкулеза у представителей русской этнической группы Челябинской области
Перечень графического материала: таблицы, диаграммы, рисунки
Дата выдачи задания " " 2008 года
Руководитель___________________
Студент________________________
![]() Содержание
Содержание
Введение
I. Обзор литературы
1.1 Физиологическая роль главного комплекса гистосовместимости человека
1.1.1 Строение системы HLA
1.1.2 Характеристика генов и антигенов HLA II класса
1.1.3 Многообразие функций системы HLA
1.1.4 Роль HLA в реализации иммунного ответа
1.1.5 Полиморфизм системы HLA и ее этнические различия
1.2 Механизмы ассоциации HLA с заболеваниями
1.3 Общее представление о туберкулезе и его ассоциация с различными генетическими факторами
1.3.1 Иммунный ответ при туберкулезе
1.4 Ассоциация HLA II класса с микобактериозами в русской популяции
II. Собственные исследования
2.1 Материалы и методы исследования
2.1.1 Контингент обследуемых лиц
2.1.2 Иммуногенетическое типирование
2.1.3 Методы статистической обработки
2.2 Результаты собственных исследований
2.2.1 Анализ распределения частот генов и антигенов HLAII класса у больных туберкулезом представителей русской национальности, проживающих в Челябинской области
2.2.2 Анализ распределения частот генов и антигенов HLAII класса в популяции русских Челябинской области
2.2.3 Сравнительный анализ частот встречаемости антигенов HLAII класса у больных туберкулезом и здоровых представителей русской этнической группы Челябинской области
2.2.4 Анализ трехлокусных гаплотипов HLA у больных туберкулезом русской этнической группы Челябинской области
2.2.5 Сравнительный анализ частот встречаемости трехлокусных гаплотипов HLA у больных туберкулезом и здоровых представителей русской этнической группы Челябинской области
Заключение
Выводы
Список использованных источников
Приложения
Список сокращений
HLA - человеческий лейкоцитарный антиген
МНС - главный комплекс гистосовместимости
PCR-SSP - полимеразная цепная реакция с аллель-специфическими праймерами
NK - натуральные киллеры
АПК – антиген-представляющие клетки
TNF - фактор некроза опухолей
ПЦР - полимеразная цепная реакция
MBL- белок, связывающий маннозу
VDR- рецептор к витамину D
Введение
Настоящая работа посвящена поиску ассоциации иммуногенетической системы HLA с развитием туберкулеза в русской этнической группе Челябинской области.
Широко распространенные популяционные исследования легли в основу бурно развивающегося направления - "HLA и болезни", т.к HLA-антигены и гены зарекомендовали себя в качестве наилучших генетических маркеров для выявления предрасположенности к ряду заболеваний. HLA-маркеры могут использоваться для прогнозирования риска развития болезней или их форм, особенностей клинического течения и эффективности терапии, для определения типа наследования и изучения генетически контролируемых механизмов патогенеза заболеваний и т.д. [6].
Накопленные сведения указывают на неоднородность ассоциаций HLA-антигенов с заболеваниями, возможность существования разных HLA-маркеров одного заболевания в различных популяциях, что отражает эволюционно сложившиеся черты их HLA-генетического профиля.
Туберкулез также относится к заболеваниям, генетические факторы предрасположенности к которому играют важную роль в возникновении и прогрессировании болезни. Понимание важной роли генетических факторов в развитии туберкулеза пришло в первую очередь из эпидемиологических и близнецовых исследований. Дальнейшие исследования, проведенные на экспериментальных животных моделях, существенно дополнили имеющуюся информацию о генетических факторах предрасположенности к заболеванию.
Данные, полученные в результате изучения распределения генов II класса главного комплекса гистосовместимости в различных этнических группах, могут быть использованы для развития направления "HLA и болезни". Таким образом, представляется целесообразным проведение исследования по обнаружению ассоциации HLA с развитием клинического туберкулеза в русской этнической группе Челябинской области.
Актуальность данных исследований в первую очередь связана с необходимостью накопления информации в направлении "HLA и болезни" в русской этнической группе Челябинской области, а также для изучения ассоциации HLA с туберкулезом.
Новизна данного исследования заключается в том, что в процессе работы впервые был проведен анализ особенностей распределения специфичностей генов HLA II класса (DRB1, DQA1 и DQB1) и их гаплотипических сочетаний на молекулярно-генетическом уровне среди пациентов с туберкулезом и здоровых людей русской этнической группы, проживающих в Челябинской области. Иммуногенетический анализ больных туберкулезом в Челябинской области проводился впервые.
Объектом настоящего исследования являлись больные туберкулезом (51 человек), находящиеся на стационарном лечении в ГУЗ "Противотуберкулезный диспансер №3", представители социально благополучных слоев населения, имеющие родственников русской национальности в трех поколениях. Группу сравнения составили 202 случайно выбранных донора Областной станции переливания крови (г. Челябинск) русской национальности.
Цель работы: Выявить наличие ассоциации генов HLAII класса (локусов DRB1, DQA1 и DQB1) и их гаплотипов с развитием туберкулеза в русской этнической группе Челябинской области.
Задачи исследования:
1. Установить частоты встречаемости генов HLAII класса и их гаплотипов в группе больных туберкулезом.
2. Провести сравнение полученных данных с результатами распределения генов HLAII класса в контрольной группе.
3. Сделать вывод о наличии положительной или отрицательной ассоциации генов HLAII класса с развитием туберкулеза у представителей русской этнической группы Челябинской области.
Теоретической базой для настоящей работы являются популяционно-иммуногенетические исследования, проведенные ранее на других популяциях, а также литературные данные об ассоциации иммуногенетической системы HLA с развитием туберкулеза.
Генотипирование аллельных вариантов HLA II класса проводилось методом полимеразной цепной реакции. Для обработки результатов применялись стандартные генетико-статистические методы.
Практическая значимость работы:
1. Данные о распределении специфичностей генов HLA II класса и их сочетаний у здоровых представителей русской этнической группы Челябинской области могут быть использованы в качестве контрольных для поиска маркеров генетической предрасположенности к развитию различных заболеваний.
2. Полученные данные могут быть теоретически использованы для практических рекомендаций при лечении туберкулеза легких.
3. Результаты исследования ассоциации генов HLAIIкласса с развитием туберкулеза могут быть использованы в качестве дополнительного диагностического критерия при выявлении группы риска по данному заболеванию.
Композиция работы определяется ее целью и задачами и отражает основные этапы исследования.
I. Обзор литературы
1.1 Физиологическая роль главного комплекса гистосовместимости человека
Регуляция иммунного ответа является одной из основных физиологических функций организма. Эта функция принадлежит генам главного комплекса гистосовместимости человека. При этом следует принять во внимание, что само это название отражает скорее историю открытия данной генетической системы, чем ее основную функцию. Дело в том, что история открытия первых продуктов генов главного комплекса гистосовместимости человека, называемых антигенами HLA (от Human leucocyte antigens), связана именно с появлением и развитием трансплантационной иммунологии, когда возникла необходимость подбора тканесовместимых пар донор и реципиент. Сегодня же известно, что роль системы HLA в отторжении трансплантата является лишь одной из частных физиологических функций этой системы, а основная же ее функция - это регуляция иммунного ответа. В 80-х годах даже дискутировался вопрос о переименовании системы HLA в главный комплекс генов иммунного ответа человека, но, учитывая, что старое историческое название давно укоренилось среди исследователей, решено было не менять его [35].
1.1.1 Строение системы HLA
По современным представлениям система HLA, обеспечивая регуляцию иммунного ответа, осуществляет такие важнейшие физиологические функции, как взаимодействие всех иммунокомпетентных клеток организма, распознавание своих и чужеродных, в том числе измененных собственных, клеток, запуск и реализацию иммунного ответа и, в целом, обеспечивает выживание человека как вида в условиях экзогенной и эндогенной агрессии [29].
Все многообразие указанных функций обеспечивается строением главного комплекса гистосовместимости [13].
Система HLA, открытая более 40 лет назад, по-прежнему остается одной из самых сложных, наиболее хорошо изученных и вместе с тем загадочных генетических структур в геноме человека. Так, если еще в 1987 г. расстояние между его условными границами оценивалось в 2000 кб, то на сегодняшний день оно расширено более чем в 2 раза, причем протяженность отдельных его элементов - генных кластеров - колеблется в широких пределах в зависимости от HLA-гаплотипа [23].
Комплекс генов HLA (главного комплекса гистосовместимости человека) компактно расположен на коротком плече 6-й аутосомной хромосомы, занимает 3500 kb (тысяч пар оснований) и содержит более 220 генов.
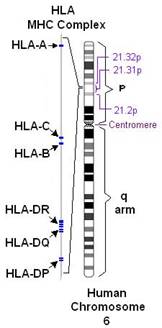
Рисунок 1. Расположение комплекса генов HLA на 6 хромосоме человека.
Гены главного комплекса гистосовместимости человека подразделяются на области A, B, С, D и обозначаются как HLA-А, HLA-В HLA-С и HLA-D, которые достаточно полиморфны и имеют варианты (аллели) и подварианты.
В последние годы в пределах I класса системы HLA открыты новые (неклассические) локусы - E, F, G, H (псевдоген, экспрессируется на уровне РНК), J (псевдоген), для большинства из которых пока не выявлено наличие аллельного полиморфизма и не определены биологические функции. Гены MIC A и MIC B расположены рядом с генами MHC I класса, но отличаются от них экзон-интронной организацией. Молекулы MIC A экспрессируются в основном на фибробластах и эпителиальных клетках. Тем не менее, предполагают, что цепь молекулы MIC A сходна с цепями молекул MHC I класса и способна связывать пептиды и другие короткие лиганды.
Между генами HLA I и II классов расположены гены, кодирующие молекулы III класса. Среди них выделяют суперсемейства C (факторы комплемента, которые вовлечены в процессы элиминации чужеродных антигенов) и G (функции окончательно не выяснены, но предполагается, что продукты экспрессии отдельных генов данного семейства участвуют в процессе созревания лейкоцитов).
На основании исследования главных комплексов гистосовместимости различных видов организмов был сделан вывод о том, что этот генный комплекс расширялся за счет дупликации, что в свою очередь давало определенные преимущества организмам с более полиморфной системой HLA в процессе эволюции. Однако вопрос о том, какие причины привели к подавлению экспрессии ранее функционировавших генов в составе МНС, остается открытым [35].
1.1.2 Характеристика генов и антигенов HLA II класса
Молекулы HLA II класса кодируются генными локусами DR, DP и DQ и имеют распространение преимущественно на макрофагах, В-лимфоцитах, активированных Т-клетках (CD4) и участвуют в гуморальном ответе. HLA-DR кодируется генами на хромосоме 6 в области 6p21.31. HLA-DR часто вовлечен в ассоциации с аутоиммунными болезнями, является протективным или ассоциированным с рядом заболеваний вместе с DQ. В случае инфекции пептид (типа энтеротоксина стафилококка) связывается с молекулой DR и DQ и представляется нескольким из очень многих TCR T-хелпера. Эти лимфоциты тогда связываются с антигенами на поверхности B-клеток, стимулируя быстрое увеличение B-клеток.
Молекулы HLA II класса - это мембранные гликопротеины, состоящие из двух цепей (α и β), каждая из которых содержит по два домена. Цепи молекул II класса очень сходны между собой. На аминокислотном уровне наиболее полиморфными являются внешние альфа-1 и бета-1 домены, которые образуют пептид-связывающую бороздку и отвечают за презентацию антигенов.
Трехмерная структура молекулы II класса была установлена в 1993 году Брауном на основе рентгеноструктурного анализа. Антигенсвязывающую бороздку молекул II класса образуют наиболее экспонированные внешние домены, α-1 и β-1. Дно бороздки сформировано b-складчатой структурой, состоящей из восьми антипараллельных участков, а стенки образованы a-спиралями (рисунок 2). Отличие антигенсвязывающей бороздки молекул II класса от таковой молекул I класса заключается в том, что в молекулах II класса она образована двумя разными цепями. Домены, образующие бороздку, и в особенности первый домен b-цепи, чрезвычайно полиморфны. Области полиморфизма собраны в несколько дискретных гипервариабельных участков (например, в b-цепи HLA-DQ они соответствуют аминокислотным остаткам в положениях 52-58, 70-77, 84-90).
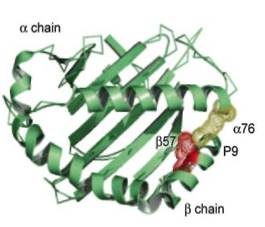
Рисунок 2. Трехмерное строение молекулы HLAII класса
Предполагают, что одни из этих участков расположены в пределах бороздки в положении, оптимальном для связывания антигена, а другие образуют детерминанты, вовлекаемые во взаимодействие с Т-клеточными рецепторами. В частности, было показано, что некоторые аминокислотные замены в полиморфных участках молекул HLA могут влиять на структурные изменения, значимые для образования "карманов", в которых связываются определенные презентируемые пептиды. Эти изменения могут даже полностью нарушать правильное связывание конкретного пептида, препятствуя его успешной презентации, а также влиять на корректное физиологическое распознавание комплекса "пептид-молекула HLA" Т-клетками. На основании полученных данных предположено, что стабильное взаимодействие молекул MHC II класса с пептидами зависит также и от плотности сети водородных связей между аминокислотными остатками альфа-спиралей молекулы MHC и связываемым пептидом. С другой стороны, установлено, что с CD4-рецепторами могут взаимодействовать и неполиморфные участки вторых доменов молекулы HLA (в данном случае HLA-DR). Таким образом, структура генов и молекул HLA обусловлена их биологическим предназначением [36].
Первичная функция HLA-II класса - представление чужеродного антигена иммунной системе для образования или подавления ответа Th, что, в конечном счете, ведет к производству антител против того же самого антигенного пептида. HLA DR найден на АПК (макрофаги, B-клетки и дендритные клетки). На поверхности клеток во время возбуждения увеличивается число антигенов DR, поэтому он является также маркером для иммунного возбуждения.
Существует чрезвычайно высокий уровень аллельного разнообразия в HLA DRB1, он стоит на втором месте после HLA-B. Эти 2 полиморфные области определяют все разнообразие последовательностей HLA в пределах человеческого генома. Это означает, что HLA-DRB1 развивается намного быстрее, чем все другие геномные локусы. Большая часть разновидностей в HLA DRB1 происходит в положениях контакта пептида в кармане, в результате многие из аллелей изменяют способ, которым DR связывает лиганды пептида и изменяет репертуар, который каждый рецептор может связать. Это свидетельствует о большом количестве изменений функционирующих в природе, и поэтому находится под действием отбора. В HLA области гены находятся в гетерозиготном состоянии или под давлением отбора, хотя некоторые аллели, возможно, находятся под положительным или отрицательным отбором.
HLA-DR и DQ локусы, вероятно, связаны с самым большим числом различных болезней относительно любых других локусов. Это происходит из-за сложной природы резистентности и высокого полиморфизма в этих локусах. Большинство этих болезней имеет низкую частоту, некоторые, подобно диабету 1 типа и целиакии являются редкими, но не исключениями.
Поскольку аллели DR находятся в неравновесном сцеплении с HLA-DQ локусами, отдельные изучения часто не могут определить точную ассоциацию с болезнью. В таких случаях используют трехлокусный гаплотип DR-DQA-DQB.
1.1.3 Многообразие функций системы HLA
Помимо того, что система HLA осуществляет регуляцию иммунного ответа на его начальных и продуктивных этапах, она также обеспечивает и такой "терминальный" этап регуляции, как апоптоз различных типов антиген-презентирующих клеток. При этом следует отметить, что этот эффект касается как профессиональных антиген-презентирующих клеток (макрофаги и CD34+ -клетки, дифференцировавшиеся из моноцитов в культуре клеток, и дендритные клетки), так и В-лимфоцитов.
Так, коллективом исследователей, работающих под руководством проф. D. Сharron, в самые последние годы было установлено, что блокирующее воздействие моноклонального антитела L243 на молекулы HLA-DR, экспрессированные на различных типах антиген-презентирующих клеток, блокирует их апоптоз, определяемый с помощью FITC-меченного аннексина V. Во всех указанных клетках после блокировки молекул DR отмечалось значительное снижение апоптоза. Авторы считают, что регуляция апоптоза дифференцирующихся антиген-представляющих клеток осуществляется через молекулы HLA-DR, и это может явиться решающим механизмом для ограничения их жизни.
Данные в целом свидетельствуют в пользу ключевой физиологической роли молекул HLA-DR в регуляции апоптоза всех типов антиген-презентующих клеток. Последнее, по сути, является регуляцией одного из важнейших этапов развития иммунного ответа и ещё раз свидетельствует в пользу того, что при современном уровне знаний о физиологической роли генов HLA-DR, можно считать, что именно в действительности являются генами иммунного ответа человека.
Генам главного комплекса гистосовместимости принадлежит ещё ряд важнейших физиологических функций. Описанию одной из них (генетическому контролю качества иммунного ответа) была посвящена работа, опубликованная в Российском физиологическом журнале им. И.М. Сеченова. Речь идет об ассоциированном с системой HLA контроле активности различных субпопуляций иммунокомпетентных клеток, что в свою очередь существенным образом сказывается на конечном уровне, т.е. на качестве иммунного ответа человека. При этом, естественно, следует помнить, что эта функция является "вторичной" и реализуется только в случае, если организм человека генетически способен отвечать на данный агент. Предпосылкой развития данного направления можно считать предположение, выдвинутое W. Bodmer и J. Bodmer еще в 1978 г., о том, что на формирование HLA-профиля европеоидной популяции в значительной степени оказали влияние имевшие место в средние века эпидемии таких заболеваний, как чума, оспа, холера и т.д. В результате этого среди выживших оказался значительный процент людей с определенными HLA-генотипами, в первую очередь с генотипом HLA-A1, - В8, - DR3. Этот генотип, как предположил W. Bodmer, обеспечивает более высокую резистентность к инфекционным заболеваниям и является на сегодняшний день генетическим маркером европеоидной популяции. Следует отметить, что это предположение было подтверждено на примере недавних вспышек брюшного тифа в Суринаме, когда среди выживших европеоидов значительный процент составили лица с гаплотипом HLA-A1, - В8, - DR3. Одновременно с этим W. Bodmer высказал справедливое предположение, что реализация этого эффекта могла быть связана только с ассоциациями между конкретными HLA-специфичностями и HLA-гаплотипами и иммунным ответом. Учитывая тот факт, что с одними и теми же гаплотипами HLA оказалась связана устойчивость к самым различным инфекционным агентам, логично было предположить, что подобного рода ассоциация с HLA может быть связана не только с самой генетически обусловленной отвечаемостью к конкретному инфекционному агенту, но и с теми звеньями иммунного ответа, которые принимают участие в его реализации, т.е. в конечном эффекте.
8-09-2015, 20:04