43 Если исходный субстрат — полисахарид, образуются 3 молекулы АТФ на 1 молекулу сброженной глюкозы.
Возникнув как первый, далекий от совершенства энергетический процесс, гомоферментативное молочнокислое брожение не было потом отброшено в процессе эволюции. Наоборот, оно закрепилось и существует сейчас в виде гликолиза у подавляющего большинства прокариот, дрожжей, грибов, а также у высших животных и растений, но только как первый этап более совершенного энергетического процесса, сформировавшегося в результате последующего развития способов получения энергии живыми организмами. Чем объясняется такая судьба гомоферментативного молочнокислого брожения? Вероятно, оказалось выгодным использовать его в качестве первого подготовительного этапа по следующим причинам: 1) высокая энергетическая эффективность (не путать с энергетическим выходом процесса!); 2) простота механизмов получения энергии; 3) перестройка исходного субстрата в форму, метаболически удобную для последующих превращений.
ГОМОФЕРМЕНТАТИВНЫЕ МОЛОЧНОКИСЛЫЕ БАКТЕРИИ
Гомоферментативное молочнокислое брожение, в основе которого лежит гликолитический путь разложения глюкозы, является единственным способом получения энергии для группы эубактерий, которые при сбраживании углеводов превращают в молочную кислоту от 85 до 90% сахара среды. Бактерии, входящие в данную группу, морфологически различны. Это кокки, относящиеся к родам Streptococcus и Pediococcus , а также длинные или короткие палочки из рода Lactobacillus . Последний подразделяется на три подрода. Бактерии, включенные в два из них (Thermobacterium, Streptobacterium ), также осуществляют гомоферментативное молочнокислое брожение. Все бактерии этой группы положительно окрашиваются по Граму, не образуют спор, неподвижны. Группа весьма гетерогенна в отношении нуклеотидного состава ДНК: молярное содержание ГЦ-пар оснований колеблется от 32 до 51%. Значительные колебания по этому признаку характерны и для бактерий, объединенных в роды и даже подроды.
Лактатдегидрогеназа, катализирующая превращение пирувата в лактат, стереоспецифична. У разных видов она содержится в виде определенных оптических изомеров; в зависимости от этого бактерии продуцируют D - или L -форму молочной кислоты. Те из них, которые образуют смесь D - и L -форм. содержат или две формы фермента, различающиеся стереоспецифичностью, или лактатрацемазу. Некоторые признаки, характерные для эубактерий, осуществляющих гомоферментативное молочнокислое брожение, представлены в табл. 15.
Таблица 15. Характеристика таксономических групп гомоферментативных молочнокислых бактерий
| Род и подрод бактерий | Морфология и особенности деления клеток | Молярное содержание ГЦ в ДНК, % | Конфигурация молочной кислоты | Наиболее распространенные виды |
| Род Streptococcus | сферические или овальные клетки; делятся в одной плоскости, в результате образуются пары или цепочки клеток | 33—44 | D | S. faecalis S. lactis |
| Род Pediococcus | кокки; делятся в двух плоскостях, в результате образуются тетрады клеток | 33—44 | DL | P. cerevisiae |
| Род Lactobacillus
Подрод Thermobacterium Подрод Streptobacterium* |
палочки; делятся в одной плоскости, образуют пары или цепочки клеток | 35—51 32—46 |
L D D DL DL L |
L. delbruckii L. bulgaricus L. lactis L. jensenii L. plantarum L. casei |
* Виды, относящиеся к этому подроду, расщепляют пентозы по окислительному пентозофосфатному пути, осуществляя гетероферментативное молочнокислое брожение. Поэтому они не являются облигатно гомоферментативными молочнокислыми бактериями.
У этой группы эубактерий молекулярный кислород не включается в энергетический метаболизм, но они способны расти в присутствии O2 , т. е. являются аэротолерантными анаэробами44 . В их клетках в значительном количестве содержатся флавиновыe ферменты, с помощью которых происходит восстановление молекулярного кислорода до H2 O2 . Из-за неспособности молочнокислых бактерий синтезировать гемовую группу у них отсутствует каталаза — фермент, катализирующий разложение перекиси водорода, поэтому последняя может накапливаться в клетке. Существующие механизмы защиты от молекулярного кислорода и его производных у этой группы эубактерий изложены в гл. 15.
44 Некоторые авторы представителей рода Lactobacillus относят к микроаэрофилам (см. сноску на с. 127).
Особенностями конструктивного метаболизма гомоферментативных молочнокислых бактерий являются слабо развитые биосинтетические способности, что выражается в большой зависимости их роста от наличия в питательной среде готовых органических веществ (аминокислоты, витамины группы В, пурины, пиримидины). В качестве источника углерода молочнокислые бактерии используют лактозу (молочный сахар) или мальтозу (растительный сахар, образующийся при гидролизе крахмала). Могут они также использовать некоторые пентозы, сахароспирты и органические кислоты. Из всех известных непатогенных прокариот молочнокислые бактерии отличаются наибольшей требовательностью к субстрату. Зависимость этих бактерий от наличия готовых органических веществ среды указывает на примитивность в целом их конструктивного метаболизма.
Молочнокислые бактерии распространены там, где они могут обеспечить свои высокие потребности в питательных веществах и где имеются большие количества углеводов, переработка которых дает им необходимую для роста энергию. Их много в молоке и молочных продуктах, на поверхности растений и в местах разложения растительных остатков; обнаружены они в пищеварительном тракте и на слизистых оболочках животных и человека.
Молочнокислым бактериям принадлежит главная роль в осуществлении ряда процессов, используемых с давних времен для получения различных кисломолочных продуктов, в процессах соления и квашения овощей, силосования кормов. Кефир — продукт совместной деятельности молочнокислых бактерий и дрожжей. Известно много национальных кисломолочных продуктов (кумыс, йогурт и др.), для приготовления которых используют кобылье, верблюжье, овечье, козье молоко, а в качестве закваски — естественно возникшие и сохраняемые комплексы молочнокислых бактерий и дрожжей. Молочнокислые бактерии играют также большую роль в процессе приготовления сыров и сливочного масла. Первый этап производства сыров (створаживание белков молока) осуществляется молочнокислыми бактериями.
Скисание сливок, необходимое для получения сливочного масла, также вызывают бактерии рода Streptococcus . Помимо молочной кислоты некоторые из них образуют ацетоин и диацетил, придающие сливочному маслу характерный запах и вкус Субстратом служит лимонная кислота, содержание которой в молоке может достигать 1 г/л. Реакции, ведущие к образованию этих веществ, начинаются с расщепления лимонной кислоты:
лимонная кислота ® уксусная кислота + щавелевоуксусная кислота
Уксусная кислота выделяется в среду, а щавелевоуксусная кислота (ЩУК) декарбоксилируется, что приводит к образованию пирувата:
ЩУК ® пировиноградная кислота + CO2 (1).
Дальнейшее метаболизирование пирувата осуществляется по трем различным путям: часть молекул восстанавливается до молочной кислоты; другая часть подвергается декарбоксилированию, приводящему к возникновению разных C2 -интермедиатов (ацетил-КоА и "активный" ацетальдегид) и взаимодействию между ними, заканчивающемуся синтезом молекулы диацетила. Восстановление последнего приводит к образованию ацетоина:
| CH3 -CO-CO-CH3 | + | НАД-H2 | ® | CH3 -CHOH-CO-CH3 | + | НАД+ | (2). |
| диацетил | ацетоин |
Эта последовательность реакций не связана с получением клеткой энергии. Смысл ее, возможно, в дополнительном своеобразном решении "акцепторной проблемы", так как, во-первых, образование пирувата в реакции 1 не сопровождается синтезом НАД-H2 , и, во-вторых, синтез ацетоина из диацетила (реакция 2) требует дополнительных молекул НАД-H2 .
Использующие мальтозу молочнокислые бактерии участвуют в квашении овощей. В мелко нарезанные овощи добавляют 2 — 3% соли и создают условия, исключающие свободный доступ воздуха. Начинается спонтанное молочнокислое брожение. Аналогичный процесс протекает при силосовании кормов. Предназначенная для силосования растительная масса плотно загружается в силосные башни или ямы. Чтобы повысить питательные свойства среды, добавляют мелассу, а с целью создания более благоприятных условий для молочнокислых бактерий растительную массу подкисляют. В этих условиях также протекает спонтанное молочнокислое брожение.
СПИРТОВОЕ БРОЖЕНИЕ
Выше мы разобрали наиболее простой способ решения донор-акцепторной проблемы, который реализуется в виде молочнокислого брожения у группы гомоферментативных молочнокислых бактерий. Дальнейшие поиски на путях эволюции привели к формированию других метаболических возможностей для решения этой проблемы. Одна из них заключается в том, что из пировиноградной кислоты в результате ее окислительного декарбоксилирования образуется, ацетальдегид, который становится конечным акцептором водорода. В итоге из 1 молекулы гексозы образуются 2 молекулы этилового спирта и 2 молекулы углекислоты. Процесс получил название спиртового брожения. Спиртовое брожение распространено среди прокариотных (различные облигатно и факультативно анаэробные эубактерии) и эукариотных (дрожжи) форм. В анаэробных условиях у высших растений также отмечено накопление этилового спирта.
Процесс спиртового брожения, осуществляемый дрожжами, до последней реакции идет по тому же пути, что и описанный выше процесс молочнокислого брожения, но последняя реакция заменена двумя другими ферментативными реакциями. Сначала пируват с помощью пируватдекарбоксилазы, ключевого фермента спиртового брожения, декарбоксилируется до ацетальдегида и CO2 :
CH3 -CO-COOH ® CH3 -COH + CO2 .
Особенность реакции заключается в ее полной необратимости. Образовавшийся ацетальдегид восстанавливается до этанола с участием НАД+ -зависимой алкогольдегидрогеназы:
CH3 -COH + НАД-H2 ® CH3 -CH2 OH + НАД+
Донором водорода служат 3-ФГА (как и в случае молочнокислого брожения).
Процесс спиртового брожения суммарно можно выразить следующим уравнением:
C6 H12 O6 + 2ФН + 2АДФ ® 2CH3 -CH2 OH + 2CO2 + 2АТФ +2H2 O.
Как видно из уравнения, с точки зрения энергетического выхода оба процесса (гомоферментативное молочнокислое и спиртовое брожение) одинаковы. В обоих случаях сбраживание 1 молекулы глюкозы приводит к образованию 2 молекул АТФ. Процессы различаются природой конечных акцепторов электронов. Кроме того, если при гомоферментативном молочнокислом брожении образовавшаяся молочная кислота в целом по степени окисленности-восстановленности не отличается от молекулы гексозы (имеет место лишь внутримолекулярное перераспределение окисленности и восстановленности отдельных углеродных атомов, входящих в ее молекулу), то в случае спиртового брожения происходит межмолекулярное размежевание на восстановленные (этиловый спирт) и окисленные (CO2 ) молекулы.
Спиртовое брожение, осуществляемое дрожжами, интересно тем, что на нем впервые были сделаны открытия, имеющие принципиальное значение. Именно при изучении спиртового брожения Л. Пастер доказал, что оно является процессом, связанным с жизнедеятельностью определенных микроорганизмов — дрожжей. Л. Пастер открыл, что в условиях свободного доступа кислорода воздуха процесс спиртового брожения ингибируется и активируется дыхание. Это явление получило название "эффекта Пастера". "Эффект Пастера" есть результат определенного взаимодействия между различными энергетическими путями, существующими у дрожжей. Одним из проявлений такого взаимодействия является конкуренция за АДФ и неорганический фосфат между процессами субстратного фосфорилирования гликолитического пути и окислительного фосфорилирования в дыхательной цепи.
В 1897 г. братья Г. и Э. Бухнеры (Н. Buchner, E. Buchner) опубликовали первое сообщение о возможности осуществления спиртового брожения вне клетки. Оказалось, что бесклеточные экстракты дрожжей превращают углеводы в этанол. Это послужило отправным пунктом для детального изучения химизма процесса. Впервые было показано включение неорганического фосфора в этот процесс и роль фосфорилированных соединений (Л. А. Иванов, 1905; A. Harden, W. Young, 1905). Установлена природа отдельных реакций, катализирующих их ферментов, промежуточных продуктов метаболизма, коферментов, энергетических взаимопревращений. В 1933г. Г. Эмбден и О.Мейергоф предложили полную схему спиртового брожения. Наконец, работы К. Нойберга (К. Neuberg) по изучению механизма спиртового брожения привели к установлению еще одной важной особенности метаболизма низших форм жизни — его чрезвычайной гибкости.
К. Нойберг обнаружил, что в зависимости от условий процесс спиртового брожения может идти с образованием продуктов, которые в норме не образуются. Если к дрожжам, сбраживающим глюкозу, добавить бисульфит, то основным продуктом брожения будет глицерин. Оказалось, что бисульфит образует комплекс с ацетальдегидом, и последний не может больше функционировать как акцептор электронов:
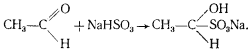
Следствием этого является передача электронов от НАД-H2 на фосфодиоксиацетон, восстановление его до 3-фосфоглицерина и дефосфорилирование последнего, приводящее к образованию глицерина. Кроме глицерина в среде происходит накопление ацетальдегида (в комплексе с бисульфитом), этанола и CO2 , но образование последних двух продуктов заметно подавлено. Когда брожение идет в присутствии бисульфита, энергетический выход процесса в два раза меньше по сравнению с нормальным спиртовым брожением, поскольку одна триоза не подвергается окислению, а восстанавливается до молекулы глицерина.
Спиртовое брожение протекает обычно при pH 3–6. Если его проводить в щелочной среде, например в присутствии NaHCO3 , также происходит накопление в сбраживаемом растворе глицерина. Оказалось, что в щелочных условиях ацетальдегид не может акцептировать электроны, поскольку в этих условиях он участвует в реакции дисмутации с образованием уксусной кислоты и этилового спирта. Акцептором электронов, как и в предыдущем случае, служит фосфодиоксиацетон. Процесс брожения в щелочной среде можно представить в виде следующего уравнения:
| 2C6 H12 O6 | ® | 2C3 H8 O3 | + | CH3 -COOH | + | CH3 -CH2 OH | + | 2CO2 . |
| глюкоза | глицерин | уксусная кислота | этиловый спирт |
МИКРООРГАНИЗМЫ, ОСУЩЕСТВЛЯЮЩИЕ СПИРТОВОЕ БРОЖЕНИЕ
Накопление этилового спирта в среде в анаэробных условиях наблюдается у разных групп эубактерий и группы эукариотных микроорганизмов — дрожжей.
Эубактерии
Способность осуществлять в анаэробных условиях спиртовое брожение по пути, описанному в предыдущем разделе, присуща некоторым эубактериям, принадлежащим к разным таксономическим группам, например Sarcina ventriculi, Erwinia amylovora .
S. ventriculi относится к группе грамположительных анаэробных кокков. Клетки неподвижные; делятся в трех плоскостях, поэтому в культуре часто образуют пакеты, состоящие из 64 и более клеток. Веществом, связывающим клетки в пакетах, служит целлюлоза. Описана способность образовывать эндоспоры. Аэротолерантный анаэроб. Единственный способ получения энергии — сбраживание Сахаров. Потребность в питательных веществах довольно высока (многочисленные аминокислоты и ряд витаминов).
E. amylovora относится к группе энтеробактерий. Это грамотрицательные подвижные палочки. Особенностью вида является его патогенность для растений. Факультативный анаэроб. В аэробных условиях получает энергию в процессе дыхания.
Помимо этилового спирта и CO2 в качестве продуктов брожения S. ventriculi в среде накапливается уксусная кислота и выделяется молекулярный водород, у E. amylovora накапливается молочная кислота. Разнообразие конечных продуктов у этих бактерий связано с тем, что пируват, образующийся при сбраживании глюкозы по гликолитическому пути, далее может метаболизироваться различно: восстанавливаться до молочной кислоты; подвергаться декарбоксилированию и последующему восстановлению, как описано в предыдущем разделе; подвергаться ферментативному расщеплению, приводящему к образованию ацетата и др.
У многих клостридиев и энтеробактерий среди продуктов брожения обнаруживают этиловый спирт, но путь его образования отличен от описанного в предыдущем разделе. Сбраживание сахаров до пировиноградной кислоты происходит по гликолитическому пути, дальнейшее же превращение пирувата идет не через пируватдекарбоксилазу. У названных групп бактерий пируват подвергается расщеплению, приводящему к образованию ацетил-КоА. Реакция катализируется пируватдегидрогеназой. Ацетил-КоА затем восстанавливается до ацетальдегида:
CH3 -CO~S-КоА + НАД-H2 ® CH3 -COH + НАД+ + КоА-SH,
а последний — до этанола.
Гетероферментативные молочнокислые бактерии накапливают в среде спирт, метаболизируя глюкозу по окислительному пентозофосфатному пути. В результате ряда ферментативных превращений образуется ацетилфосфат, восстановление которого в два этапа приводит к появлению молекулы этилового спирта.
Наконец, у бактерий Zymomonas mobilis с неясным систематическим положением, используемых в Мексике для получения национального спиртного напитка "пульке", разложение глюкозы до пировиноградной кислоты идет по пути Энтнера — Дудорова. Дальнейшее превращение пирувата происходит с участием пируватдекарбоксилазы и алкогольдегидрогеназы. Выход продуктов брожения такой же, как при спиртовом брожении по гликолитическому пути: по 2 молекулы спирта и CO2 на 1 молекулу сброженной глюкозы, но энергетический выход в два раза ниже, чем при гликолизе: всего 1 молекула АТФ на 1 молекулу сброженной глюкозы.
Z. mobilis — грамотрицательные подвижные бактерии, имеющие форму коротких палочек. Характеризуются высокими биосинтетическими способностями. Анаэробы, единственный способ получения энергии для которых — спиртовое брожение. Однако эти бактерии способны расти в присутствии молекулярного кислорода. Последний в этом случае используется для окисления части этанола до уксусной кислоты в соответствии с уравнением:
1 глюкоза + 1O2 ® 1 этиловый спирт + 1 уксусная кислота + 1,7CO2 + 0,2 молочная кислота
Таким образом, молекулярный кислород существенно не меняет характера энергетического метаболизма Z. mobilis. В клетках бактерии обнаружены фрагменты ЦТК, цитохромы b , c , a2 , каталаза. Наиболее вероятным представляется, что предки Z. mobilis — аэробы. Способ получения энергии за счет спиртового брожения — более позднее приспособление к условиям обитания.
Эукариоты
Основными продуцентами этилового спирта, имеющими широкое практическое применение, являются дрожжи — одноклеточные эукариотные микроорганизмы, принадлежащие к разным классам высших грибов. Наиболее распространенный способ размножения дрожжей — почкование. Дрожжи — аэробы со сформированным аппаратом дыхания, но в анаэробных условиях осуществляют спиртовое брожение по пути, рассмотренному в предыдущем разделе, т. е. получают энергию за счет субстратного фосфорилирования. Конструктивный метаболизм дрожжей основан на их хорошо развитых биосинтетических способностях. Есть виды дрожжей, развивающиеся на простых синтетических средах; эти дрожжи способны синтезировать все необходимые им сложные органические соединения. Существуют виды,. нуждающиеся в определенных витаминах группы В. Добавление к питательной среде веществ, содержащих комплекс витаминов, аминокислот, сахаров приводит, как правило, к заметному стимулированию роста дрожжей.
Ряд отраслей промышленности основан на жизнедеятельности дрожжей (виноделие, производство спирта, пивоварение, хлебопекарное производство). Сырьем для производства спирта с использованием дрожжей служат углеводы растительного происхождения (картофель, злаки), отходы пищевой (мелассы) и целлюлозно-бумажной (щелока) промышленности, различные сельскохозяйственные отходы, а также гидролизаты древесины. Сбраживание дрожжами виноградного сока лежит в основе виноделия; сбраживание пивного сусла, приготовленного из проросших зерен ячменя, специальными пивными дрожжами — в основе пивоварения.
О путях образования этилового спирта
Изложенные данные позволяют составить определенное представление о том, насколько широко распространено образование этилового спирта среди разных групп эубактерий и насколько различны метаболические пути, ведущие к его синтезу. Из этого следует, что накопление в культуральной среде этилового спирта само по себе не может служить указанием на место процесса, приводящего к его образованию, в эволюции. Этиловый спирт у эубактерий может быть одним из конечных продуктов как эволюционно более ранних (гликолиз), так и более поздних (окислительный пентозофосфатный цикл, путь Энтнера — Дудорова) катаболических процессов. До сих пор среди эубактерий не обнаружены организмы, сохранившие черты примитивности энергетического и конструктивного метаболизма, у которых спиртовое брожение служило бы единственным способом получения энергии. Тот факт, что в самом "классическом" виде спиртовое брожение проявляется у дрожжей, форм эукариотных, не может, как нам кажется, ставить под сомнение его место в эволюции анаэробных энергетических процессов.
ПРОПИОНОВОКИСЛОЕ БРОЖЕНИЕ
Из рассмотренных двух типов брожения видно, что ключевым
29-04-2015, 01:57