| ЗАРО | n | Нейтрофіли, Г/л | НСТТс, % |
ФІН, % |
ФЧН, мікр./фаг. |
ІБЦН, % |
БЦЗН, Г/л |
|
| Переактивація | 10 | X±m d±m |
2,65±0,32 -0,72±0,22 Іа |
14,7±2,9 -0,20±0,240 |
58,5±4,8 -1,35±0,37Іб |
7,2±0,4 -0,43±0,210 |
19,8±1,8 -1,82±0,10ІІа |
3,80±0,60 -1,26±0,09 Іа |
| Підвищена активація НРР | 28 | X±m d±m |
3,21±0,16 -0,33±0,110 |
17,4±1,3 -0,64±0,09 Іа |
55,5±1,9 -1,58±0,14 Іб |
6,0±0,3 -1,05±0,15 Іа |
20,6±0,9 -1,78±0,05 ІІа |
4,12±0,36 -1,21±0,06 Іа |
Тренування НРР |
4 | X±m d±m |
5,24±0,52 1,06±0,36 Іа |
20,3±3,2 -0,89±0,12 Іа |
52,3±1,9 -1,83±0,14 |
6,8±0,7 -0,67±0,40 Іа |
18,6±2,1 -1,89±0,12 ІІа |
6,43±1,08 -0,85±0,17 Іа |
| Спокійна активація НРР | 9 | X±m d±m |
4,88±0,31 0,81±0,21 Іа |
11,1±1,3 -0,07±0,180 |
61,1±2,0 -1,15±0,15 Іа |
6,3±0,5 -0,89±0,28 Іа |
27,0±2,1 -1,42±0,12 Іб |
8,16±1,04 -0,57±0,16 Іа |
Тренування ВРР |
5 | X±m d±m |
3,07±0,03 -0,43±0,020 |
8,2±0,1 0,34±0,030 |
63,5±0,2 -0,97±0,02 Іа |
8,5±0,2 0,27±0,120 |
38,1±0,1 -0,81±0,01 Іа |
9,95±0,31 -0,29±0,050 |
| Підвищена активація ВРР | 16 | X±m d±m |
3,18±0,23 -0,36±0,160 |
9,7±0,8 0,15±0,120 |
64,9±1,7 -0,86±0,13 Іа |
7,8±0,3 -0,10±0,180 |
34,3±1,3 -1,01±0,07 Іа |
8,44±0,68 -0,53±0,11Іа |
| Спокійна активація ВРР | 6 | X±m d±m |
3,30±0,33 -0,27±0,220 |
9,3±0,8 0,16±0,170 |
74,8±2,1 -0,10±0,160 |
8,0±0,2 0±0,090 |
51,3±2,1 -0,07±0,120 |
13,96±2,03 0,34±0,320 |
| Донори | 20 | X±m | 3,70±0,31 | 9,7±1,2 | 76,1±2,5 | 8,0±0,3 | 52,6±2,2 | 11,7±1,2 |
Примітки: 1. X±m - актуальні середні величини та їх похибки.
2. d±m - сигмальні відхилення показників від норми та їх похибки.
3. Відзначено ступінь відхилення показників від норми.
При визначенні вмісту субпопуляцій лімфоцитів (табл. 4) виявлялась така ж тенденція, а саме - дизбаланс в складі різних субпопуляцій більш виражений за дизгармонійних ЗАРО.
Таблиця 4. Характеристика стану клітинної ланки імунітету (відносні показники) при різних ЗАРО
| ЗАРО | CD3-Л, % |
CD4-Л, % |
0-лімф., % |
РБТЛ, % |
CD8-Л, % |
CD16-Л, % |
ПКА, % |
АЗЦ, % |
| Переактивація | 48,9±1,7 -0,95±0,29Іа |
36,6±2,1 0,15±0,280 |
17,8±1,8 -1,42±0,04Іб |
47,2±2,2 -1,42±0,18 Іб |
21,6±1,0 -0,82±0,24 Іа |
2,5±0,3 -1,31±0,03 Іб |
3,9±0,4 -1,16±0,02 Іа |
8,9±1,3 -1,48±0,08 Іб |
| Підвищена активація НРР | 49,5±1,0 -0,86±0,17 Іа |
32,2±1,4 -0,45±0,200 |
17,4±1,4 -1,41±0,04 Іб |
46,2±1,7 -1,50±0,13 Іб |
23,1±0,7 -0,44±0,190 |
5,5±0,5 -0,99±0,06 Іа |
9,3±1,2 -0,92±0,05 Іа |
22,2±1,6 -0,64±0,10 Іа |
Тренування НРР |
51,0±2,2 -0,60±0,370 |
27,5±3,4 -1,09±0,47Іа |
25,1±4,1 -1,56±0,04 Іб |
46,8±2,8 -1,46±0,22 Іб |
25,8±1,4 0,25±0,370 |
3,0±0,5 -1,25±0,05 Іа |
4,5±0,4 -1,13±0,02 Іа |
9,8±0,8 -1,43±0,05 Іб |
| Спокійна активація НРР | 50,9±1,4 -0,63±0,25 Іа |
37,8±1,9 0,31±0,250 |
9,7±1,7 -1,02±0,13 Іа |
49,6±2,4 -1,24±0,19 Іа |
22,0±1,0 -0,72±0,26 Іа |
9,9±1,0 -0,53±0,11 Іа |
19,1±2,7 -0,47±0,120 |
33,1±1,5 0,05±0,090 |
Тренування ВРР |
49,6±2,7 -0,85±0,42 Іа |
38,8±2,2 0,45±0,290 |
16,3±3,9 -1,38±0,09 Іб |
42,5±2,5 -1,79±0,19ІІа |
21,0±1,0 -0,97±0,25 Іа |
3,3±0,5 -1,23±0,05 Іа |
5,4±0,7 -1,09±0,03 Іа |
10,2±1,0 -1,40±0,06 Іб |
| Підвищена активація ВРР | 49,4±1,6 -0,88±0,28 Іа |
34,9±1,4 -0,08±0,190 |
14,4±1,9 -1,30±0,07 Іб |
41,6±2,7 -1,87±0,21 ІІа |
22,1±0,9 -0,68±0,23 Іа |
6,9±0,9 -0,84±0,10 Іа |
12,0±2,0 -0,79±0,09 Іа |
25,4±2,5 -0,44±0,160 |
| Спокійна активація ВРР | 54,3±0,8 -0,04±0,140 |
35,6±0,8 0,02±0,110 |
4,5±1,2 -0,02±0,180 |
61,8±3,4 -0,27±0,270 |
24,3±0,4 -0,13±0,110 |
14,7±0,7 -0,03±0,080 |
28,2±1,8 -0,05±0,080 |
32,3±1,0 0±0,070 |
| Донори | 54,5±1,3 | 35,5±1,6 | 4,5±1,3 | 65,2±2,8 | 24,8±0,9 | 14,9±2,1 | 29,4±4,9 | 32,3±3,5 |
Функціональні імунні показники, такі як антитілозалежна цитотоксична активність та природна цитотоксична активність, бласттрансформуюча здатність Т-клітин були також суттєво знижені порівняно з нормою. Було найбільше зниження за ЗАРО переактивації та тренування, як гармонійного так і дизгармонійного, де виявлено зниження ПКА до 3-5% при нормі 28-30%, а АЗЦ - до 8-10% при нормі 33-35%. Пригнічення бласттрансформації було на 20-30% і виявлено за всіх типів ЗАРО, крім гармонійної спокійної активації.
Показники гуморального імунітету також були змінені, але на відміну від Т-лімфоцитів, було виявлено практично 1,2-1,4 разове збільшення кількості В-лімфоцитів, за винятком гармонійної ЗАРО тренування, де було відмічено 2-кратне зниження рівня СD19-клітин, що рівнялось 0,21±0,01 Г/л при нормі 0,4±0,02 Г/л.
Стосовно окремих класів імуноглобулінів було виявлено, що рівень IgG був знижений за всіх без винятку типів ЗАРО, хоча найбільше його зниження було за реакцій тренування та переактивації - до 7,0-8,5 г/л при нормі 13,0±0,6 г/л, в той час як рівні ІgА та IgМ були збільшені на 20-40% в порівнянні з нормою. Практично у всіх групах виявлене підвищення рівня циркулюючих імунних комплексів.
Таким чином, проведені дослідження виявили, що у ліквідаторів, хворих на КПН, мають місце порушення в усіх ланках імунітету, в активності щитовидної, статевих та надниркових залоз, яке взаємопов'язується з певними типами гармонійних та дизгармонійних ЗАРО. В той же час складно зробити узагальнюючий висновок, виходячи з різнонаправлених та різнокількісних змін в досліджуваних показниках.
Кластерний аналіз параметрів адаптації та імунітету. В той час як рутинний методичний підхід дозволяє лише почергово аналізувати ту чи іншу ознаку статистичної вибірки, застосування кластерного аналізу робить можливим одночасне врахування усіх ознак. Врахування усієї сукупності ознак осіб, взятих в їх взаємозв’язку і зумовленості одних з них іншими дає змогу здійснити природну класифікацію, яка відображує природу речей, їх суть. Вважається, що пізнання суті об’єкта зводиться до виявлення тих його якісних властивостей, котрі власне і визначають даний об’єкт, відрізняють його від інших (Aldenderfer M.S., Blashfield R.K., 1985).
Кластеризація параметрів імунітету реалізована ітеративним методом k-means. В даному методі об’єкт відносять до того класу, евклідова віддаль до якого мінімальна. Головний принцип структурного підходу до виділення однорідних груп полягає в тім, що об’єкти одного класу близькі, а різних – віддалені. Іншими словами, кластер (образ) – таке нагромадження точок в n-мірному геометричному просторі, в якому середня міжточкова віддаль менша ніж середня віддаль від даних точок до решти.
Із врахованих 45 параметрів семи типів ЗАРО створено 10 кластерів. До першого кластеру увійшло 3 параметри, евклідові віддалі до яких від респективного центру кластера знаходяться в інтервалі 0,15ч0,37; до другого - 3 (0,27ч0,49); до третього - 5 (0,22ч0,47); до четвертого - 7 (0,14ч0,61); до п'ятого - 5 (0,13ч0,30); до шостого - 8 (0,13ч0,26); до сьомого - 2 (0,42); до восьмого - 3 (0,46ч1,05); до дев'ятого - 4 (0,18ч0,42) і до десятого - 5 (0,11ч0,31). Отже, кластеризація проведена цілком коректно. Це підтверджується також дисперсійним аналізом. Долі різних типів ЗАРО в дисперсії, пояснюваній розподілом параметрів на кластери, знаходяться в діапазоні 94,8ч87,5%, за винятком гармонійної реакції спокійної активації, яка складає лише 61,5%. Коефіцієнти канонікальної кореляції: 0,974ч0,935 і 0,784 відповідно.
У перший кластер включено абсолютний вміст Т-гелперів/індукторів, теофілінчутливих Т-лімфоцитів та В-лімфоцитів. Другий кластер об'єднує абсолютний вміст популяції Т-лімфоцитів і їх теофілінчутливої субпопуляції, а також фагоцитарний індекс моноцитів-макрофагів Третій кластер включає в себе відносний вміст теофілінчутливих Т-лімфоцитів і Т-кіллерів, а також абсолютний вміст пан-нейтрофілів, долю в них нейтрофілів, які експресують FcIgG-рецептори, і абсолютний вміст фагоцитуючих нейтрофілів. Четвертий кластер об'єднує відносний вміст В-лімфоцитів і Т-гелперів/індукторів - з одного боку, фагоцитарні числа макрофагів і мікрофагів та параметри кисеньзалежних механізмів бактерицидності останніх - з іншого боку. У п'ятий кластер увійшли параметри кіллерної ланки імунітету, а також концентрація IgG і абсолютний вміст "активних" Т-лімфоцитів. Найчисленніший за кількістю параметрів шостий кластер об'єднує CD3-лімфоцити і їх ТФР-субпопуляцію, комплемент, вміст моноцитів і їх фагоцитарну ємність, а також мікробну ємність і бактерицидну здатність нейтрофілів. У сьомому кластері виявилося лише два параметри В-ланки - рівень IgM та крупномолекулярних ЦІК, а у восьмому кластері - ще три (IgA та ЦІК середніх і дрібних розмірів). Дев'ятий кластер об'єднує лізоцим і функціональні параметри Т-ланки (Еа-РУЛ, РБТЛ, 0-лімфоцити). Нарешті, параметри десятого кластера стосуються виключно фагоцитозу нейтрофілів.
Виявилось, що лише гармонійна ЗАРО спокійної активації супроводжується нормальними параметрами імунітету і неспецифічного захисту, тоді як стосовно інших станів адаптації дійти однозначних висновків на основі проведеного аналізу неможливо. Це спонукало продовжити аналіз з використанням інших методичних підходів.
Факторний аналіз інформаційного поля парамерів адаптації та імунітету. Графічним методом Scree-test Cattel - за моментом виходу графіка величин власних чисел на плато - відібрано для подальшого аналізу вісім ГК, кумулятивна доля котрих у поясненні сумарної дисперсії поля 68 параметрів складає 76,7%.
Перша ГК, за означенням, відтворює максимальну долю варіабельності інформаційного поля - 23,0%. Вона пов'язана із 16 параметрами, в тому числі суттєво (r>0,70) - із 10. Всі вони стосуються виключно здатності нейтрофілів фіксувати, поглинати і знешкоджувати мікроби. Окрім того, тут виявлено показник інтенсивності фагоцитозу моноцитів та активність комплемента - одного із опсонінів. Із переліченими параметрами антибактеріального захисту поєднані індекс адаптації Поповича та мінералокортикоїдна активність. Друга ГК пояснює 14,5% дисперсії і пов'язана із 9 параметрами, які стосуються абсолютного вмісту загальних лімфоцитів та їх Т- і В-популяцій, а також лейкоцитів в цілому. Третя ГК поглинає 10,5% дисперсії і стосується, з одного боку, вмісту нейтрофілів та В-лімфоцитів, а з іншого - глюкокортикоїдної функції. Натомість четверта ГК (8,1% дисперсії) пов'язана з параметрами кіллерної ланки імунітету та андрогенною і тиреоїдною функціями адаптивних залоз. П'ята ГК (6,7% дисперсії) стосується так званих імунорегуляторних індексів, разом з котрими виявилися фагоцитарний індекс моноцитів та індекс напруження еозинофілів лейкограми. Решта параметрів моноцитів-макрофагів, а також індекс кіллінгу нейтрофілів-мікрофагів об'єднані у шостій ГК, яка пояснює 5,8% дисперсії. Сьома ГК поглинає 4,7% дисперсії, об'єднуючи, по-перше, параметри відносного вмісту та активності субпопуляцій Т-лімфоцитів, по-друге, гуморальних факторів антибактеріального захисту, джерелами яких є В-лімфоцити (Igg M i G) та нейтрофіли і моноцити (лізоцим). Нарешті, восьма ГК (3,4% дисперсії) об'єднує решту параметрів В-ланки, а також індекси напруження двох компонент лейкограми.
Отже, понад 3/4 дисперсії інформації про стан пристосувально-захисних механізмів обстеженого контингенту, яка міститься у 68 параметрах-змінних, конденсується у восьми головних компонентах, тобто може бути пояснений обмеженим числом іх лінійних комбінацій.
Дискримінантний аналіз функціональних параметрів, які характеризують тип ЗАРО. З метою виявлення параметрів лейкограми, адаптивних залоз та імунітету, констелляція яких характерна для кожного із семи типів ЗАРО, наявне інформаційне поле із 68 параметрів було піддано дискримінантному аналізу (метод forward stepwise). Для включення в модель відібрано 26 параметрів (в порядку зниження критерію Л): вміст в крові лімфоцитів, тироксину, індекс бактерицидності нейтрофілів, Na/K-коефіцієнт плазми, антитілазалежна цитотоксичність, вміст нейтрофілів, реакція бласттрансформації лімфоцитів, вміст еозинофілів і моноцитів, екскреція з сечею 17-OKС та 17-KС, вміст CD16-лімфоцитів, індекс напруження еозинофілів, вміст теофілінрезистентних і "активних" Т-лімфоцитів, фагоцитарне число моноцитів, бактерицидна здатність мікрофагів, вміст лейкоцитів, CD4-лімфоцитів, теофілінчутливих Т-лімфоцитів, В- і 0-лімфоцитів, індекс напруження моноцитів, активність лізоциму, індекс напруження лейкоцитів та вміст паличкоядерних нейтрофілів.
За сукупністю відібраних змінних стани пристосувально-захисних систем осіб усіх семи груп суттєво відрізняються між собою: міжгрупові віддалі Mahalanobis складають 4,7-19,0 (р<10-4 ).
Перша дискримінантна функція характеризує відносний вміст лімфоцитів в лейкоцитограмі, тобто кардинальний типоутворюючий параметр ЗАРО. Друга функція найтісніше інверсно пов'язана із параметрами бактерицидності нейтрофілів і лізоцимом сироватки та вмістом натуральних кіллерів - з одного боку, та мінералокортикоідної (інверсно) і глюкокортикоїдної (прямо) функціями - з іншого. Третя функція теж пов'язана із індексом бактерицидності, але прямо, що асоціюється з інверсною кореляцією з мінералокортикоїдною функцією та прямою - з тиреоїдною і глюкокортикоїдною функціями. Четверта функція, як і перша, негативно корелює із лімфоцитами лейкоцитограми, разом з тим, має місце зв'язок також з моноцитами, індексом їх напруження і фагоцитарним числом; проте найтісніше вона пов'язана з тиреоїдною функцією, в меншій мірі - з андрогенною, що асоціюється із антитілазалежною цитотоксичністю. Останній параметр, разом з нейтрофільзом, в найбільшій мірі характеризують п'яту функцію, котра, своєю чергою, прямо корелює із тиреоїдною і андрогенною функціями. Натомість остання, шоста функція не корелює суттєво з жодним параметром.
Сума добутків нестандартизованих коефіцієнтів на значення дискримінантних змінних разом із константою дають значення дискримінантної функції для кожної особи. Значення дискримінантних функцій визначають точку в їх просторі. Дані візуалізовані на площині перших трьох функцій (радикалів), котрі в сумі містять 87,4% дискримінантних можливостей (рис. 1).
Отже, розбіжності між типами ЗАРО вичерпно пояснюються 26 параметрами, з-поміж яких 8 стосуються лейкоцитограми, 4 - ендокринних залоз, 5 - фагоцитарної ланки імунітету, 6 - Т-клітинної, 2 - кіллерної та 1 - В-клітинної ланок імунітету. Інформація, що міститься в цих параметрах, може бути сконденсована у шести, а по суті - у трьох функціях-радикалах.
Відібрані параметри можуть бути використані для ідентифікації того чи іншого типу ЗАРО. Ця мета дискримінантного аналізу реалізується з допомогою класифікуючих (дискримінантних) функцій - особливих лінійних комбінацій для кожної групи, які максимізують розбіжності між групами і мінімізують дисперсію всередині груп.
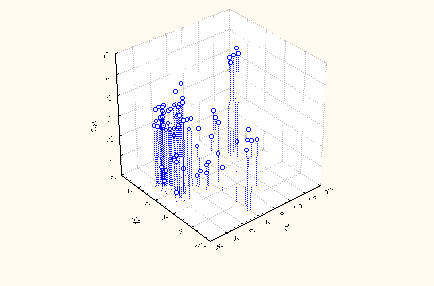
Рис. 1. Діаграма розсіювання нестандартизованих канонікальних величин перших трьох коренів інформаційного поля обстеженого контингенту осіб.
Канонікальний аналіз кореляційних зв'язків між параметрами адаптації та імунітету. На першому етапі канонікального кореляційного аналізу було з'ясовано зв'язки між констелляцією чотирьох маркерів функціонального стану адаптивних залоз, з одного боку, та набором десяти параметрів лейкограми периферійної крові - з іншого. Коефіцієнт канонікальної кореляції (r*) між першими радикалами, добутими із параметрів функціонального стану адаптивних залоз і лейкоцитограми периферійної крові, складає 0,961 (ч2 =217; Л Prime=0,044; p<10-6 ). Отже, функціональний стан головних адаптивних залоз на 92% детермінує лейкоцитограму. Іншими словами, остання є адекватним відображенням ЗАРО.
Наступний етап було присвячено дослідженню зв'язків ІАП як кількісного виразу ЗАРО із функціональним станом адаптивних залоз та лейкограмою. Величина r* складає 0,871 (ч2 =65; Л Prime=0,39; p<10-6 ). Радикал даного інформаційного поля корелює із моноцитами (r=-0,72); ІН моноцитів (r=0,66); лімфоцитами (r=0,42); 17-КС (r=-0,41) та ІН еозинофілів (r=0,29) і детермінує величину ІАП на 61%.
Рівняння регресії для обчислення ІАП має вигляд:
ІАП = 0,454 + 0,474*М - 1,733*ІНМ + 0,06*(17-КС) - 0,043*Лф
R2 = 0,448; F(4,73) = 14,9; p<10-5
Далі досліджено зв'язки ІАП з вісьмома параметрами В-ланки імунітету. Виявлено його інверсну кореляцію із вмістом в сироватці ЦІК середніх (r=-0,53) і дрібних (r=-0,49) розмірів та IgA (r=-0,40). У підсумку величина ІАП визначається станом В-ланки імунітету на 51% (r*=0,714; ч2 =51; Л Prime=0,49; p<10-4 ).
З-поміж 20 параметрів кіллерної та Т-ланок імунітету суттєві зв'язки з ІАП констатовано стосовно рівня натуральних кіллерів (r=0,48), зумовленої ними природної кіллерної активності (r=0,46), а також антитілазалежної цитотоксичності (r=0,34) - з одного боку, і вмістом "активних" Т-лімфоцитів (r=0,32) та 0-лімфоцитів (r=-0,35) - з іншого. Міра детермінації ІАП (r*=0,665; ч2 =38,6; Л Prime=0,56; p=0,008).
З-поміж 23 параметрів фагоцитарної ланки імунітету ІАП значуще корелює із 18, в тому числі тісно із індексами кіллінгу (r=0,76) і бактерицидності (r=0,84) нейтрофілів та їх бактерицидною здатністю (r=0,71); посередньо - із мікробною ємністю моноцитів (r=0,60), їх фагоцитарним числом (r=0,55) та кількістю моноцитів (r=0,51), активністю лізоциму сироватки (r=0,54), фагоцитарним індексом нейтрофілів (r=0,51), вмістом нейтрофілів, які експресують C3b -рецептори (r=0,51); слабо - із вмістом нейтрофілів, які експресують FcIgG-рецептори (r=0,49), активністю в нейтрофілах мієлопероксидази (r=0,50), лізосомально-катіонного тесту (r=0,50) та тестів з нітросинім тетразолієм, причому з активованим - прямо (r=0,445), а із спонтанним - інверсно (r=-0,47), а також похідним від двох останніх показником функціонального резерву мікрофагів (r=0,49). До переліку параметрів із слабким зв'язком із ІАП належать також вміст в крові макрофагів (r=0,47), фагоцитарне (мікробне) число нейтрофілів (r=0,44) та їх мікробна ємність (r=0,38).
ІАП детермінується даною захисною системою на 78% (r*=0,881; ч2 =96; Л Prime=0,22; p<10-6 ).
Базуючись на отриманих результатах канонікального аналізу, на заключному етапі було сформовано констелляцію із параметрів кожної із систем, які найтісніше корелюють із величиною ІАП, а саме (в порядку зменшення модуля коефіцієнта кореляції із радикалом ІАП): індекс бактерицидності нейтрофілів (r=-0,96), їх бактерицидна здатність (r=-0,81), мікробна ємність моноцитів (r=-0,69), відносний вміст моноцитів (r=-0,65), активність лізоциму (r=-0,62), вміст ЦІК середніх розмірів (r=0,61), індекс напруження моноцитів (r=0,60), вміст натуральних кіллерів (r=-0,55), IgA (r=0,46), 0-лімфоцитів (r=0,41), пан-лімфоцитів (r=0,38), Еа-РУЛ (r=-0,36) та екскреція 17-КС (r=-0,36).
У підсумку величина r* склала 0,871 (ч2 =98; Л Prime=0,24; p<10-6 ). Іншими словами, величина ІАП на 76% детермінується констелляцією 13 параметрів пристосувально-захисних систем. Обмежившись вісьмома із них, ми склали рівняння множинної регресії для обчислення ІАП:
ІАП=0,02-0,119*ІБЦН-0,034*БЦЗН+0,331*МЄМ-0,008*Ліз-2,42*ЦІКсер+1,78*ЦІКдр+0,092*CD16+0,05*ФЧН
R2 =0,744; F(8,69) =25; p<10-5
Отже, індекс адаптації Поповича адекватно відображує стан пристосувально-захисних систем організму за різних типів його загальних адаптаційних реакцій.
Інший аспект канонікального аналізу стосується зв'язків 15 параметрів ЗАРО (4 - маркерів стану адаптивних залоз, 10 - елементів лейкоцитограми і індексів їх напруження, а також ІАП) - з одного боку, та параметрів окремих ланок імунітету - з іншого боку. Стосовно В-ланки r* складає 0,986 (ч2 =413; Л Prime=0,002; p<10-6 ); об'єднаної Т- і кіллерної ланок: 0,988 (ч2 =702; Л Prime=10-5 ; p<10-6 ); фагоцитарної ланки імунітету: 0,997 (ч2 = 1187; p<10-6 ).
Отже, параметри ЗАРО тісно корелюють із параметрами антибактеріальних захисних систем, детермінуючи їх на 97-99%, відображує патогенетичні зв'язки між станом адаптації і резистентності організму.
Ефективність стандартного бальнеотерапевтичного комплексу курорту Трускавець. Лікування хворих покращило, як і слід було очікувати, їх стан здоров'я, а також ЗАРО, що в цілому узгоджується з даними багатьох авторів, які також констатували позитивний вплив бальнеотерапії на стан здоров'я хворих з різною патологією (Алексєєв О.І. та ін., 1994,1995; Бажан К.В., 1998; Бульба А.Я., 2002; Прокопович Л.Н., 2006).
В проведених нами спостереженнях за хворими на калькульозний пієлонефрит ліквідаторів було встановлено, що по закінченню курсу лікування на курорті Трускавець оздоровчий ефект в значній мірі залежав від клінічного синдрому, а саме найбільший був ефект при больовому синдромі, який під впливом бальнеотерапії знижався в 3,8 рази за шкалою Наrrington у хворих на активну форму КПН. У хворих на активну форму КПН з дизуричним та сечовим синдромом спостерігалось зменшення клінічних проявів за шкалою Наrrington в 1,5-2 рази, тоді як у хворих з диспептичним та енцефалопатичним синдромами клінічні прояви захворювання не зменшувались після лікування на курорті Трускавець. У хворих з латентною формою КПН та хворих в фазі ремісії, де клінічні прояви були слабо виражені, під час лікування виявилось незначне покращення по шкалі Наrrington, це складало 10-25% від величини індекса при поступленні на лікування, тобто цей показник вкладається у відомі клінічні критерії „без змін" та „незначне покращення".
Пострадіаційний енцефалопатичний синдром має максимальну часткову вираженість і він в найменшій мірі зменшувався після лікування на курорті Трускавець, тоді як дизуричний та сечовий синдром в значний мірі (в два і більше разів) зменшувався.
Сумарно виявлено значне покращення у 25% хворих, покращення - у 70% і лише у 5% - незначне покращення, що свідчить про позитивний вплив на клінічну картину захворювання. Клінічне покращення підтверджується даними дослідження сечі, де виявлено зменшення лейкоцитурії та бактерійурії. Так, у хворих в активній фазі захворювання лейкоцитурія при поступленні складала 32,8*106
/л, а по закінченні лікування - всього 3,6*106
/л, тобто в 7-8 разів знижувалась лейкоцитурія. При інших формах КПН зниження було меншим, хоча і сама лейкоцитурія була незначна - від 1 до 5*106
/л. Особливо показове зменшення бактерійурії, яке при активній формі КПН складало в 10 разів - з 1065*106
до 103*106
КУО/л, тоді як в латентній формі зменшення було менш значним - з 217*106
до 12*106
КУО/л. Таким чином, під впливом бальнеологічного лікування відбувалось суттєве зменшення запального та інфекційного процесу при калькульозному пієлонефриті, що свідчить про позитивний вплив води Нафтуся на всі
8-09-2015, 19:30