Загальна закономірність збільшення амплітуди ПН при збільшенні ступеня навантаження на стегнову кістку була характерна як для контрольних, так і для дослідних щурів. Однак у тварин, які перебували в умовах гіпокінезії, значення приросту амплітуди ПН були завжди менше, ніж у контрольних. Як у контрольних, так і у дослідних щурів значення електричного потенціалу, що доводиться на одиницю навантаження, при впливах I і II ступеня збільшувалися, а III ступеня - зменшувалися. Однак середні значення А/Н у дослідних тварин при навантаженнях I і II ступеня були на 36% менше, ніж у контрольних. При навантаженнях III ступеня вірогідних розходжень в амплітуді ПН і значенні А/Н між контрольними та дослідними тваринами невстановлено.
Дослідження впливу вікових змін кісткової тканини на ПН стегнових кісток проводили на щурах віком 2 , 6 і 24 міс. Загальна тенденція до збільшення амплітуди ПН зі збільшенням навантаження на кістку була властива для тварин усіх вікових груп.
Зростаючи в експонентній залежності, ПН сягав максимальних значень при навантаженнях III ступеня. При всіх ступенях навантаження максимальні амплітуди ПН були зареєстровані у стегнових кістках 6-місячних щурів. У 2-місячних щурів ПН не перевищував 9,1-10,3-11,6% амплітуди ПН 6-місячних тварин при відповідних ступенях навантаження. У 24-місячних тварин ПН був лише 54,5-78,7-81,4% такого показника 6-місячних щурів при відповідних ступенях навантаження. Слід зазначити, що найбільші розходження амплітуди ПН у 2, 6 і 24-місячних щурів були зафіксовані при навантаженнях I та II ступеня.
Незалежно від віку щурів, при збільшенні навантажень від 30 до 50% від маси тіла значення електричного потенціалу, що доводиться на одиницю навантаження, збільшувалися, а при навантаженнях, які сягали 100% від маси тіла - знижувалися.
Максимальне значення цього показника було зареєстровано в 6-місячних щурів і сягало 6,3-7,8 мкВ/г. У 2-місячних щурів він не перевищував 0,8-0,9 мкВ/г, а у щурів 24-місячного віку він зменшувався до 2,8-4,2 мкВ/г.
Пасивні електричні властивості компактної кістки білих щурів при стереотипному функціональному навантаженні. При мультичастотному імпедансометричному тестуванні електрична ємність препаратів стегнової кістки контрольних щурів змінювалася згідно з експонентною залежністю, досягаючи мінімуму на частоті 1 МГц.
Максимальне значення реактивного опору реєстрували на частоті 10 кГц. Воно суттєво зменшувалося як на більш високих, так і на більш низьких частотах. Оскільки реактивний опір відображає стан поляризаційних процесів у біологічних тканинах, можна говорити про те, що на частоті 104 Гц вони сягали максимального ступеня виразності. Властивості частотної динаміки імпедансу й активного опору були дуже близькі. На частотах 104 ,105 Гц їх значення суттєво не змінювалися, на частотах 102 ,103 Гц вони зростали, а на частоті 106 Гц – зменшувалися.
Для встановлення зв'язку вікових змін структури та складу кістки з її ПЕВ були проведені дослідження на 49 щурах-самцях віком від 1 до 24 міс. Для стегнової кістки одномісячних щурів були характерні низькі значення імпедансу, активного та реактивного опору і високі показники ємності.
Із віком тварини імпеданс, активний і реактивний опір збільшувалися, а ємність зменшувалася. Найбільш значні зміни ПЕВ відзначалися протягом перших 3-4-х місяців життя. У дорослих щурів показники ПЕВ залишалися відносно сталими, а після 18 міс значення імпедансу, активного й реактивного опору починали поступово зменшуватися, а ємності - збільшуватися.
Маса стегнових кісток збільшувалася втричі протягом перших 3-х місяців життя, від 6 до 12 міс - сягала пікових значень, а після 18 міс починала зменшуватися, що свідчило про початок вікової інволюції кісткової тканини. Остеометричні дослідження показали, що діаметр кістковомозкового каналу в період активного росту щурів (1-6 міс) збільшувався істотніше. У тварин віком від 6 до 14 міс його показники залишалися відносно стабільними, а після 14 міс він знову починав збільшуватися. Товщина стінки діафіза в перші 3-4 міс життя щурів також зростала більше ніж удвічі. Потім темпи його приросту істотно сповільнювалися та зберігалися такими до 12-місячного віку. У щурів після 18 міс життя товщина стінки діафіза починала поступово зменшуватись. Аналіз закономірностей вікової динаміки остеометричних показників стегнових кісток щурів дозволив виділити найбільш характерні періоди їх змін. Вони відповідали віку 1-3, 6-14, 18-24 міс і практично повністю збігалися з описаними вище періодами вікових змін ПЕВ кістки. Дослідження вікової динаміки основних складових кісткового матриксу показало, що вміст мінеральних речовин істотно збільшувався у перші 3-4 міс життя щура (з 45,5±1,1 до 56,6±1,6%). Із віком мінералізація кістки продовжувала збільшуватися, але значно меншими темпами. У щурів віком від 12 до 18 міс масова частка мінеральних речовин залишалася стабільно високою (60,4±0,2%), а після 18 міс почала відзначатися чітко виражена тенденція до її зниження. У 24-місячних тварин масова частка мінеральних речовин у складі кісткового матриксу не перевищувала 53,7± 1,3%. Вікова динаміка вмісту органічних речовин наближалася до динаміки вмісту мінеральних речовин. У одномісячних щурів масова частка води у кістковій тканині сягала 35,0±1,4%, у щурів 6-місячного віку вона зменшувалася до 17,0±0,3%. У тварин, вік яких перевищував 18 міс, вміст води в кістковій тканині починав поступово збільшуватися й до 24-місячного віку сягав 26,39±1,35%. Щільність кістки також найбільше зростала в перші 3 міс життя щурів. До 18 міс вона залишалася стабільною, а потім починала поступово зменшуватися.
Кореляційний аналіз показав, що між віковими змінами показників гідратації та мінералізації компактної кістки щурів існувала значна зворотна залежність (r=-0,894; р<0,01). Показники вмісту органічних і мінеральних речовин також були пов'язані зворотною кореляційною залежністю, однак ступінь її виразності був значно меншим (r=-0,406; p<0,05). Аналіз кореляційних зв'язків показників ПЕВ і складу кістки показав наявність вірогідного прямого зв'язку між ємністю, вмістом води й органічних речовин (r=+0,858; r=+0,485) та імпедансом, активним і реактивним опором і вмістом мінеральних речовин (r=+0,766; r=+0,765; r=+0,893). Вірогідний зворотний кореляційний зв'язок був виявлений між ємністю і вмістом мінеральних речовин (r=-0,877), а також між імпедансом, активним, реактивним опором і вмістом води (r=-0,594; r=-0,592; r=-0,901) та органічних речовин (r= -0,491; r=-0,392; r=-0,304). Отримані результати свідчать про те, що вікові розходження пасивних електричних властивостей стегнової кістки щурів визначаються особливостями її складу та структури в різні періоди життєвого циклу.
Пасивні електричні властивості кістки при дозованій гіпокінезії. Після 28-добової жорсткої гіпокінезії у 67% дослідних тварин спостерігалися характерні ознаки остеопенії – низька маса й щільність кістки в порівнянні з тваринами контрольної групи. В 33% щурів ці показники не відрізнялися від контролю. Отримані результати свідчили про наявність міжіндивідуальної варіабельності та відносної "інертності" реакції компактної кістки щурів на зменшення функціонального навантаження, що узгоджується з даними інших дослідників. У кістках тварин, що мали ознаки остеопенії, відзначалося вірогідне зменшення об'ємної маси мінеральних речовин (на 15,6%) та збільшення вмісту води (на 10,6%). Порівняльний аналіз показав, що у контрольних тварин щільність сухої кістки сягала 1,95±0,09 мг/мм3 , а у тварин, що мали ознаки остеопенії - не перевищувала 1,65±0,13 мг/мм3 (р<0,05). Зменшення функціонального навантаження на кінцівки в умовах жорсткого 28-добового обмеження рухливості істотно не впливало на ріст стегнової кістки в довжину, але помітно знижувало темпи її аппозиційного росту, що призводило до зменшення маси стегнової кістки, середнього діаметра діафіза та просвіту кістковомозкового каналу на 16,6, 19,0 і 20,4% відповідно (p<0,05).
Для ПЕВ препаратів стегнових кісток, що мали ознаки остеопенії, на всьому діапазоні частот були властиві низькі величини електричного імпедансу, активного й реактивного опору і високі показники електричної ємності. Так, на частоті 104 Гц зменшення середньої величини електричного імпедансу сягало 50,1%, реактивного й активного опорів - 80,4 і 50,2% відповідно при збільшенні електричної ємності на 396,0%.
При збільшенні тривалості гіпокінезії до 45 діб у всіх дослідних щурів спостерігалися ознаки остеопенії. Маса стегнової кістки тварин була вірогідно меншою, ніж контролю (750,01±65,01 і 894,55±30,61 мг відповідно), а щільність кісткової тканини не перевищувала 1,62±0,04 мг/мм3 (р<0,05). Вміст мінеральних і органічних речовин у компактній кістці дослідних щурів вірогідно зменшувався, а вміст води - збільшувався. Якщо в контрольній групі щурів мінеральна насиченість кісткової тканини становила 1,46±0,03 мг/мм3 , то у тварин дослідної групи вона не перевищувала 1,23±0,04 мг/мм3 (p<0,001). У щурів, що перебували в умовах 45-добової гіпокінезії, відзначалася більш висока гідратація кісткової тканини порівняно зі щурами контрольної групи і порівняно з тваринами, які знаходилися в умовах жорсткої 28-добової гіпокінезії (0,43±0,01, 0,73±0,05 і 0,52±0,03 мг/мм3 відповідно). Характер і загальна спрямованість змін діаметра діафіза та кістковомозкового каналу стегнових кісток щурів після жорсткої 45-добової гіпокінезії був таким, як і у тварин після жорсткої 28-добової гіпокінезії, проте ступінь їх виразності був найбільшим, що призводило до вірогідного зменшення товщини стінки діафіза на 22,2% (р<0,05).
Гіпокінетичні порушення складу й остеометричних параметрів стегнової кістки дослідних щурів супроводжувалися характерними змінами її ПЕВ. На рис.10 представлена динаміка ємності, імпедансу, активного та реактивного опору препаратів стегнової кістки тварин контрольної й дослідної груп при проведенні мультичаcтотного імпедансометричного тестування.
Загальна спрямованість змін ПЕВ препаратів тварин після 45-добової гіпокінезії на всьому діапазоні частот була такою самою, як і у тварин після 28-добової гіпокінезії, однак ступінь їх виразності був значно більшим. На частоті 104 Гц питома ємність і реактивний опір становили 773,8 і 17,1% відповідно щодо контролю. Вірогідно, хоча не так значно, зменшувалися імпеданс і активний опір (до 26,6 і 26,4% відповідно щодо контролю).
Залежність електричних показників кістки від змін складу та фізико-хімічних властивостей основних компонентів кісткового матриксу. Для моделювання змін водно-мінерального балансу кістки в умовах гіпокінезії використаний метод демінералізації препаратів in vitro. Контролем були недемінералізовані препарати тих самих тварин.
Демінералізація препаратів кістки супроводжувалася зменшенням їхньої вихідної маси на 55% та збільшенням вмісту води на 48%. Істотно змінювалися ПЕВ препаратів. Якщо імпедансу свіжовиділених препаратів сягав 1,5 кОм, то після демінералізації він не перевищував 40 Ом. Активний опір до кінця періоду демінералізації зменшувався більше ніж у 34 рази, а реактивний опір - в 60 разів. Демінералізовані препарати втрачали залежність показників ПЕВ від частоти тестового струму. Очевидно, що в умовах гіпокінезії повної демінералізації кістки ніколи не відбувається, але і сама модель хімічної модифікації кісткової тканини лише з відомим ступенем припущення відображає реальні процеси, що відбуваються в кістці при зменшенні функціонального навантаження. Однак два факти, що випливають із результатів цієї серії досліджень, становлять інтерес і можуть бути використані для пояснення особливостей зміни пасивних електричних властивостей кістки в умовах реальної гіпокінезії: 1 - процеси демінералізації та гіпергідратації кістки є двома взаємозалежними процесами, які протікають паралельно; 2 - гіпомінералізація та гіпергідратація кістки спричинюють збільшення її електропровідності й ємності.
Для дослідження впливу змін фізико-хімічних властивостей органічного матриксу на ПЕВ кісткової тканини використаний метод дозованої термічної денатурації. Встановлено, що при нагріванні препаратів кістки від 37 до 65єС ємність, імпеданс, активний і реактивний опір істотно не змінювалися. Подальше нагрівання препаратів до 100єС супроводжувалося вірогідним збільшенням ємності на 105% і зменшенням на 47% імпедансу, активного й реактивного опору, що свідчило про розвиток денатураційних процесів в органічному матриксі .
Той факт, що зони фазових змін основних електричних показників кістки відзначалися при температурі, що перевищувала 600 С і були подібні до зон фазового переходу колагену у процесі його термічної денатурації, дає змогу говорити про наявність зв'язку між ПЕВ кістки та фізико-хімічними властивостями її органічного матриксу і, в першу чергу, колагену.
Кисневий метаболізм і пасивні електричні властивості кістки при гіпокінезії в умовах нормоксії і переривчастої нормобаричної гіпоксії. Напруження кисню (РО2 ) у литковому м'язі щурів, які перебували в умовах жорсткої 28-добової гіпокінезії в атмосферному повітрі, не перевищувала 13,5±1,7 мм рт.ст., що на 43,7% менше відповідного показника контрольних тварин. РО2 у литковому м'язі щурів, які перебували в умовах жорсткої гіпокінезії та дихали нормобаричними гіпоксичними газовими сумішами, було вірогідно вище і сягало 19,2±1,9 мм рт.ст. (p<0,05).
Споживання кисню (VО2 ) кістковою тканиною діафізарної частини стегнової кістки контрольних щурів становило 0,09±0,012 млО2 ·100г-1 ·хв-1 .
У тварин, які перебували в умовах жорсткої 28 добової гіпокінезії в атмосферному повітрі, VО2 не перевищувало 0,03±0,011 млО2 ·100г-1 ·хв-1 (p<0,05). При аналогічних режимах гіпокінезії в умовах переривчастої нормобаричної гіпоксії величини споживання кисню практично не відрізнялися від показників контрольної групи тварин. У щурів, які перебували в умовах гіпокінезії та протягом 8 год щодня дихали НГГС, маса стегнової кістки була лише на 15,5% (p<0,05) меншою, ніж у тварин контрольної групи. Діаметр діафіза та просвіту кістковомозкового каналу, а також товщина стінки діафіза не відрізнялися від відповідних показників контрольної групи тварин, хоча загальна тенденція до їх зменшення ще спостерігалася.
Аналіз складу кістки показав, що у щурів, які перебували в умовах гіпокінезії та дихали НГГС, відмінності у вмісті мінеральних речовин і води із тваринами контрольної групи не перевищували 9 і 21% відповідно (p<0,05). При цьому відзначалася характерна тенденція до збільшення вмісту органічних речовин. У щурів, що одержували НГГС на етапі преадаптації й у період дії гіпокінезії, маса та щільність кістки не відрізнялися від маси кістки тварин контрольної групи. Не було статистично достовірних відмінностей і у значеннях основних остеометричних показників, а також у вмісті мінеральних речовин і води. Однак тенденція до збільшення вмісту органічних речовин мала характер статистично значимої закономірності. Якщо вміст органічних речовин у компактній кістці тварин контрольної групи становив 0,52±0,03 мг/мм3 , то у тварин дослідної групи він сягав 0,72 ±0,02 мг/мм3 (p<0,05).
Дихання НГГС у період обмеження рухливості щурів хоча й не попереджало повністю порушень ПЕВ кісткової тканини щурів, але істотно зменшувало ступінь їх виразності. Розбіжності у значеннях ємності, імпедансу, активного і реактивного опору між дослідними та контрольними тваринами не перевищували 26,5, 30,6, 30,5 і 22,5% відповідно. Використання НГГС на етапі 14-добової преадаптації та в умовах гіпокінезії призводило до вірогідного зменшення ємності на 42,2%, p<0,05, збільшення імпедансу, активного та реактивного опору на 43,2, 43,3 і 78,5% відповідно, p<0,05. Такі зміни показників ПЕВ кістки цілком закономірно можна пов'язати зі зміною її складу й, у першу чергу, збільшенням вмісту органічних речовин, які, як це відомо, мають більшу поляризаційну здатність і меншу електропровідність.
Виходячи з результатів проведених досліджень, вплив гіпокінезії на біоелектричні властивості кістки можна представити у вигляді наступної схеми:
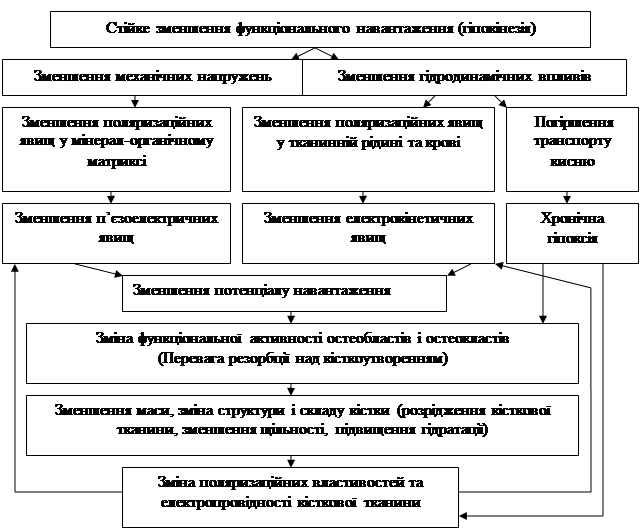

Таким чином, на підставі проведених досліджень можна стверджувати, що відсутність адекватного навантаження на кістку може бути причиною виникнення низки змін з боку клітинних і позаклітинних елементів кісткової тканини, які призводять до порушень її здатності генерувати електричні потенціали у відповідь на механічне навантаження та забезпечувати необхідний рівень біоелектричної регуляції процесів фізіологічної перебудови кістки
ВИСНОВКИ
У дисертації наведено результати досліджень впливу гіпокінезії на біоелектричні властивості стегнової кістки щурів, які отримані методом визначення електричного потенціалу навантаження на поверхні кістки та мультичастотного імпедансометричного тестування в умовах дозованого механічного навантаження й обмеження рухливості. Отримано нові результати про закономірності змін потенціалу навантаження та пасивних електричних властивостей кістки в різних режимах гіпокінезії і переривчастої нормобаричної гіпоксії, які можуть бути використані для розробки нових підходів до діагностики й корекції патології кісткової системи.
1. Електричний потенціал, який виникає на поверхні стегнової кістки щура при механічному навантаженні, зростає в експоненціальній залежності від величини прикладеного зусилля. Найбільший приріст потенціалу на одиницю навантаження забезпечують впливи, що не перевищують 50% маси тіла тварини.
2. У періоді постнатального життя щурів електричний потенціал навантаження стегнової кістки залежить від віку тварини. Амплітуда потенціалу та його приріст на одиницю навантаження найменші у щурів пубертатного періоду, у дорослих - вони сягають максимальних значень, а в періоді вікової інволюції – зменшуються.
3. У дорослих щурів після жорсткої 28-добової гіпокінезії електричний по-тенціал навантаження знижується на 20-25%. Найбільш чітко ці зміни виявляються у діапазоні навантажень, що не перевищують 50% від маси тіла тварини. Це може бути однією з причин низької ефективності профілактичного використання фізичних навантажень при розвитку вікової остеопенії та уповільненого відновлення кісткової тканини в період післядії гіпокінезії.
4. Вікові відмінності пасивних електричних властивостей стегнової кістки щурів визначаються особливостями її складу та структури в різні періоди життєвого циклу. Встановлено наявність вірогідного прямого кореляційного зв'язку між електричною ємністю та вмістом води (r=+0,858) і органічних речовин (r=+0,485), а також між імпедансом, активним і реактивним опором і вмістом мінеральних речовин (r=+0,766; r=+0,765; r=+0,893). Найбільш істотні зміни пасивних електричних властивостей кістки щурів відбуваються в періоди пубертації (1-3 міс) та інволюції (вік тварин перевищує 18 міс).
5. Жорстке тривале обмеження рухливості щурів призводить до зменшення маси, щільності та збільшення ступеня гідратації кістки. Для розгорнутої картини індукованої гіпокінезією остеопенії притаманно зменшення імпедансу, активного й реактивного опору і значне збільшення ємності кісткової тканини.
6. Критичним етапом у зміні пасивних електричних властивостей кістки при низькому функціональному навантаженні є підвищення гідратації кісткової тканини, що призводить до значного збільшення її електропровідності. Наявність
8-09-2015, 23:24